
Układ dopełniacza
ODPORNOŚĆ NIESWOISTA – CZĘŚĆ II

Co to jest układ dopełniacza?
Układem dopełniacza (komplementu) nazywamy zespół kilkudziesięciu białek
odpowiedzialnych za wrodzoną odporność humoralną, obecnych w osoczu i innych płynach
ustrojowych. W skład układu dopełniacza wchodzą również liczne białka o charakterze
regulacyjnym.
Charakterystyczną cechą układu dopełniacza jest współzależność jego składowych, a co za
tym idzie – kaskadowość jego działania.
Białka układu dopełniacza produkowane są głównie przez komórki wątroby (hepatocyty) oraz
monocyty.
Układ dopełniacza może zostać aktywowany na trzy sposoby:
Drogą klasyczną
(z udziałem przeciwciał)
Drogą lektynową
(z udziałem kolektyn)
Drogą alternatywną - properdynowa
(niezależną od białek wiążących się z patogenami)

Funkcje układu dopełniacza
Stymulacja ostrego stanu zapalnego poprzez aktywację
degranulacji komórek tucznych (rola anafilatoksyn C3a i C5a)
Chemotaktyczne wabienie neutrofilów do miejsca ataku patogenów
(rola składnika C5a)
Opsonizacja komórek patogenów (składniki C3b, iC3b, C4b) i
ułatwianie ich fagocytozy (receptory CR1 = CD35 są obecne na
monocytach, makrofagach, neutrofilach i erytrocytach)
Wspomaganie tworzenia kompleksów immunologicznych (C3b
opłaszczający patogeny, wiązany jest przez CR1 na erytrocytach, a
całość wędruje do śledziony i wątroby, gdzie zachodzi fagocytoza)
Niszczenie komórek patogennych na drodze aktywacji kompleksu
atakującego błonę (MAC)

Regulacja układu dopełniacza
Duża reaktywność poszczególnych składowych dopełniacza stanowi
niebezpieczeństwo nie tylko dla patogenów, ale i dla zdrowych komórek
organizmu. Dlatego też, mimo krótkiego czasu półtrwania, aktywność białek
dopełniacza musi być regulowana przez dodatkowe czynniki obecne w
osoczu oraz w błonach komórkowych wszystkich komórek organizmu.
Białka regulatorowe w osoczu
Białka regulatorowe w błonie
Czynnik I
(rozkłada C3b i C4b oraz
powoduje rozpad konwertaz)
Receptor CR1
(rozkłada C3b i C4b oraz
powoduje rozkład konwertaz)
Inhibitor C1
(hamuje proteazy C1r i
C1s)
MCP = CD46, błonowy kofaktor
białkowy
(wiąże C3b i C4b)
Białko wiążące C4
(wiąże C4b,
przyspiesza odłączanie C2b)
DAF, czynnik przyspieszający
rozkład
(skraca czas życia konwertaz)
Czynnik H
(wiąże C3b)
HRF = CD59, czynnik restrykcji
homologicznej
(wiąże C8 i C9, blokuje
powstawanie MAC)
Białko S
(blokuje wiązanie C5b67 do
błony komórkowej)
MIRL
(blokuje przyłączenie C9 i
powstawanie MAC)
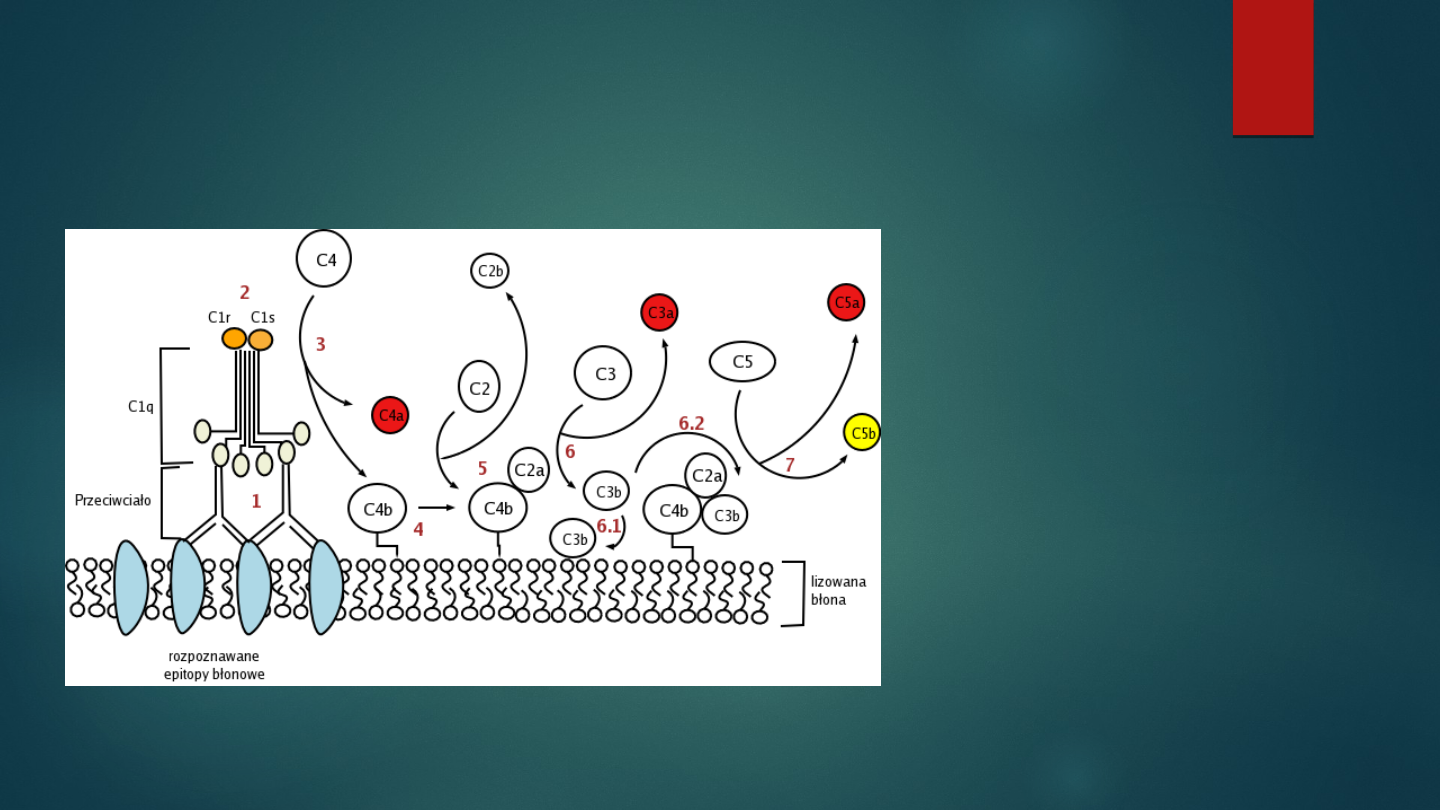
Aktywacja dopełniacza
droga klasyczna
1. Połączenie
C1
z Ab aktywacja
proteaz serynowych
C1r
i
C1s
2. Rozkład C4 na
C4a
i C4b,
przyłączenie
C4b
do błony komórkowej
3. Przyłączenie C2 do
C4b
, rozkład C2
do
C2a
i C2b – powstaje
C4b2a
–
konwertaza C3 drogi klasycznej
4. Rozkład C3 do
C3a
i
C3b
(która może
działać jako opsonina lub połączyć się z
C4b2a
tworząc
C4b2a3b
– konwertazę
C5
drogi klasycznej)
5. Rozkład C5 do
C5a
i
C5b
tworzenie
MAC

Aktywacja dopełniacza
droga lektynowa
Droga lektynowa jest zasadniczo bardzo zbliżona do
drogi klasycznej, jednak w tym wypadku w aktywacji
kaskady dopełniacza nie biorą udziału przeciwciała.
Rolę składnika C1 pełni
MBL
(białko wiążące mannozę)
lub
białka surfaktantu płucnego A i D
, które mają
zdolność wiązania się z resztami cukrowymi w błonie
komórkowej patogenów.
Związanie się MBL do błony komórkowej bakterii
powoduje zmiany konformacyjne aktywujące proteazy
serynowe
MASP1
i
MASP2
Aktywna proteaza
MASP2
ma zdolność cięcia składnika
C4 i C2, a dalsze etapy aktywacji komplementu
przebiegają jak w drodze klasycznej.
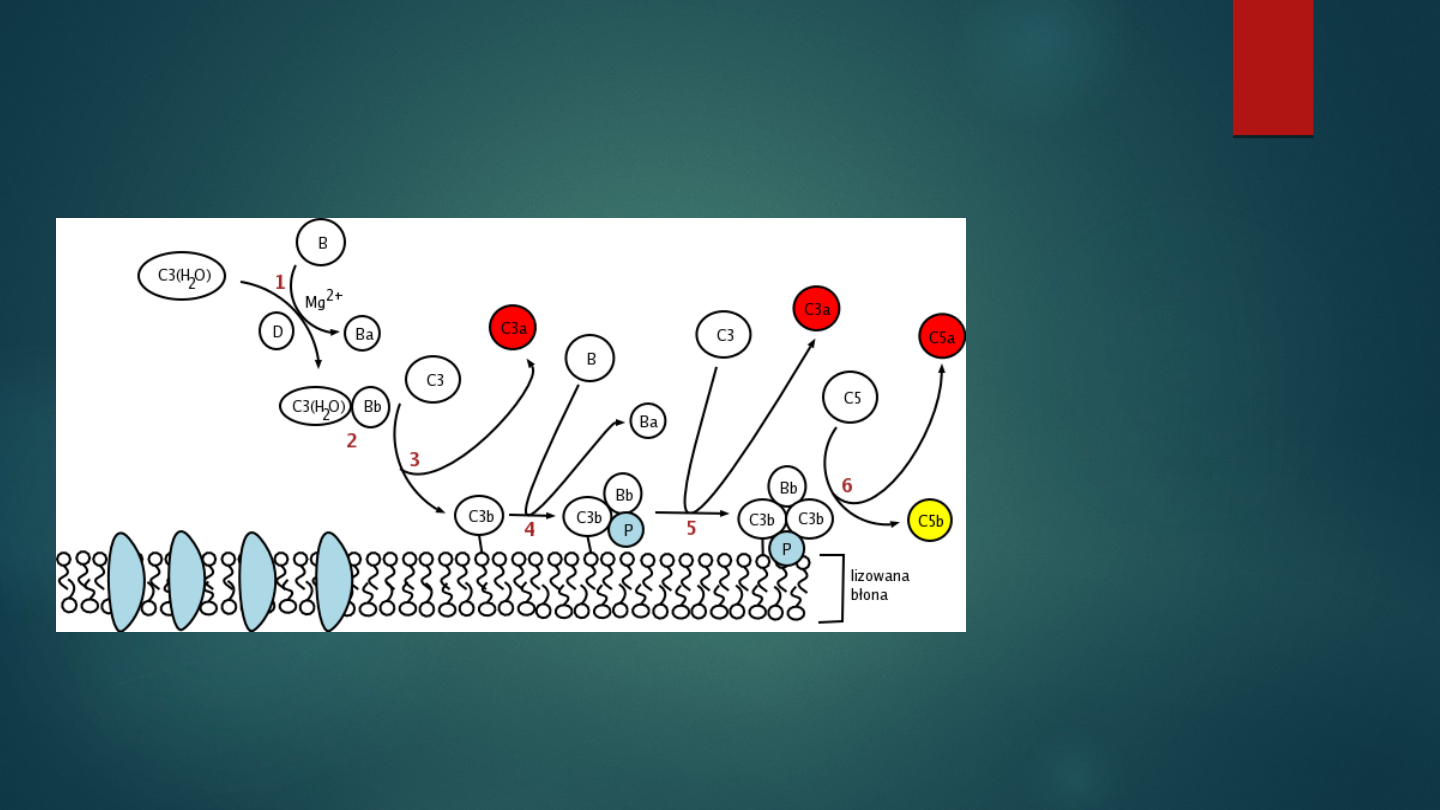
Aktywacja dopełniacza
droga alternatywna
Ważne:
konwertaza C3 drogi
alternatywnej:
C3bBb
Konwertaza C5 drogi
alternatywnej:
C3bBb3b
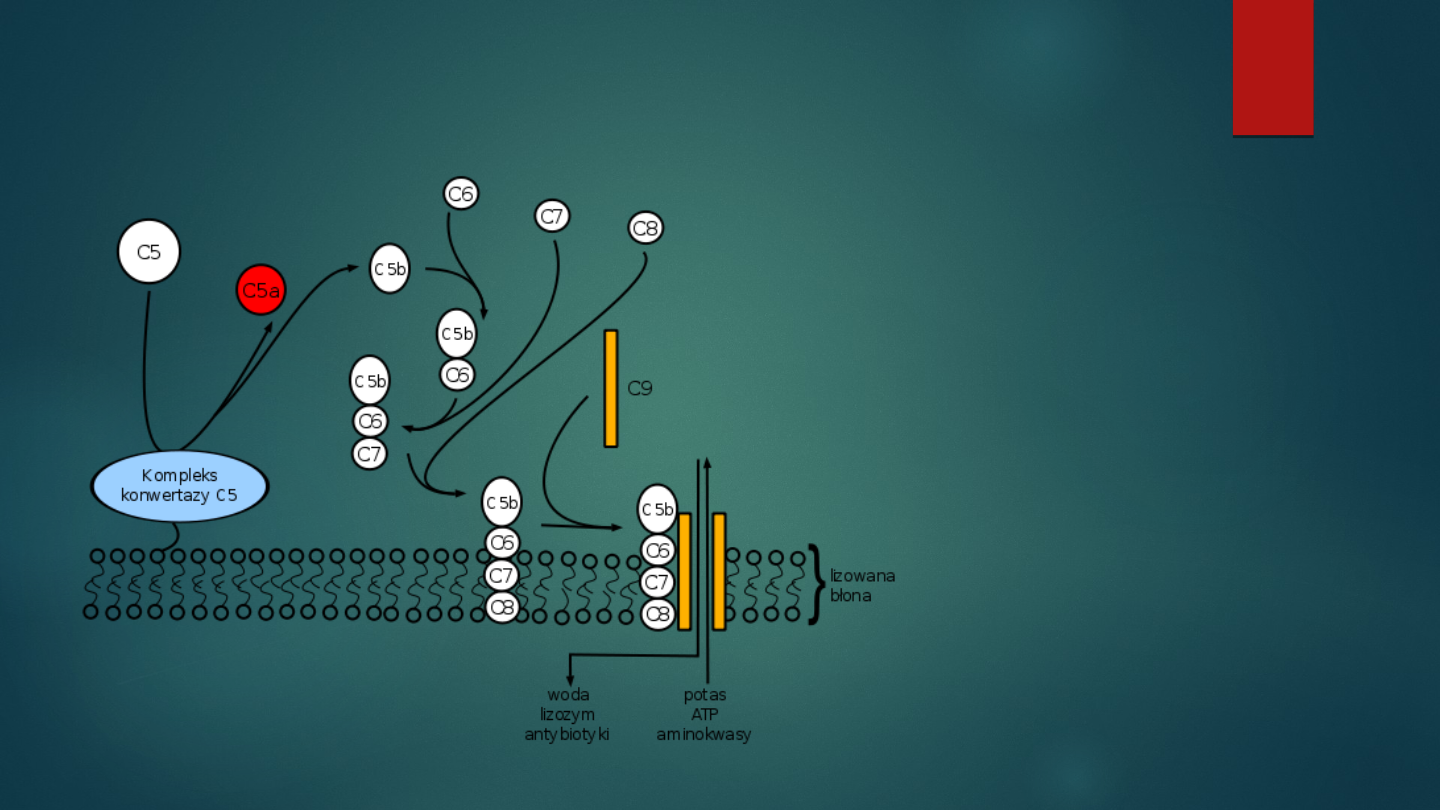
Kompleks atakujący błonę
Document Outline
- Slide 1
- Co to jest układ dopełniacza?
- Funkcje układu dopełniacza
- Regulacja układu dopełniacza
- Aktywacja dopełniacza droga klasyczna
- Aktywacja dopełniacza droga lektynowa
- Aktywacja dopełniacza droga alternatywna
- Kompleks atakujący błonę
Wyszukiwarka
Podobne podstrony:
Wykład 3, 4 układ dopełniacza
UKŁAD DOPEŁNIACZA
UKŁAD DOPEŁNIACZA
Układ dopełniacza
Uklad pokarmowy
Układ mięśniowy
układ moczowy
Układ nerwowy
oddechowy uklad
Uklad oddechowy2
T5 UKŁAD HYDRAYLICZNY PODNOSZENIA OSPRZĘT DODATKOWY
UKŁAD PŁCIOWY MĘSKI ptt
układ naczyniowy wstep
Uklad oddech wyklad
W 11 Leki działające pobudzająco na ośrodkowy układ
UKŁAD MOCZOWY WYWIADY
więcej podobnych podstron