11.05.2013
Oczos04 - BioInfoArticles
bioinfo.mol.uj.edu.pl/articles/Oczos04
1/4
Rola układu dopełniacza w mechanizmach odporności
Jadwiga Oczoś
Streszczenie: Dopełniacz (ang. complement) to układ kilkudziesięciu białek odpornościowych surowicy krwi, które po uaktywnieniu oddziałują ze sobą
w odpowiedniej kolejności. Efektem kaskadowej aktywacji jest m.in. liza drobnoustrojów chorobotwórczych. Układ dopełniacza jest istotnym
elementem wrodzonych mechanizmów odporności nieswoistej. Jednak przy zaburzonej regulacji nadmierna aktywność dopełniacza wiedzie do stanów
patologicznych, jak wstrząs anafilaktyczny, przewlekłe zapalenie. Wiązanie dopełniacza przez kompleksy immunologiczne znalazło zastosowanie w
diagnostyce kiły (odczyn Wassermanna).
Białka układu dopełniacza
Układ dopełniacza u człowieka składa się z ponad 20 rozpuszczalnych glikoprotein. Większość z nich jest wytwarzana przez hepatocyty i monocyty.
Obecność białek dopełniacza jest konstytutywna we krwi i innych płynach ustrojowych. Do białek układu dopełniacza należą:
C1 (C1q, C1r, C1s), C2, C3, C4, C5, C6, C7, C8, C9. Białka te były nazywane w kolejności odkrywania, dlatego cyfry arabskie przy literze
"C" nie oddają kolejności udziału w reakcji dopełniacza,
Czynniki B, D, H, I oraz properdyna P białka związane z alternatywną drogą aktywacji dopełniacza,
Lektyna wiążąca mannozę MBL (ang. mannose binding lectin), proteazy serynowe MASP1, MASP2 (ang. MBL associated serine proteases)
białka obecne w drodze lektynowej aktywacji dopełniacza,
inhibitor C1, C4BP (ang. C4binding protein), DAF (ang. decay accelerating factor), MCP (ang. membrane cofactor protein), CD59
(protektyna), białko S (witronektyna), receptor C1 (CR1) białka regulatorowe.
Aktywacja dopełniacza.
Aktywacja dopełniacza polega na serii enzymatycznych i nieenzymatycznych reakcji o charakterze kaskadowym. Możliwe są trzy drogi aktywacji
dopełniacza: klasyczna, alternatywna oraz lektynowa. W przypadku każdej z dróg dochodzi do utworzenia dwóch istotnych enzymów: konwertazy C3 i
konwertazy C5, które bardzo silnie wzmacniają efekt dopełniacza. Niezależnie od sposobu aktywacji końcowe etapy wszystkich trzech kaskad są
identyczne i doprowadzają do utworzenia kompleksu atakującego błonę MAC (ang. membrane attacking complex), odpowiedzialnego za lizę bakterii.
Droga klasyczna
Cechą charakterystyczną klasycznej drogi aktywacji dopełniacza jest zależność od przeciwciał. Przeciwciała zdolne do aktywacji to IgM oraz IgG
(oprócz podklasy IgG4).
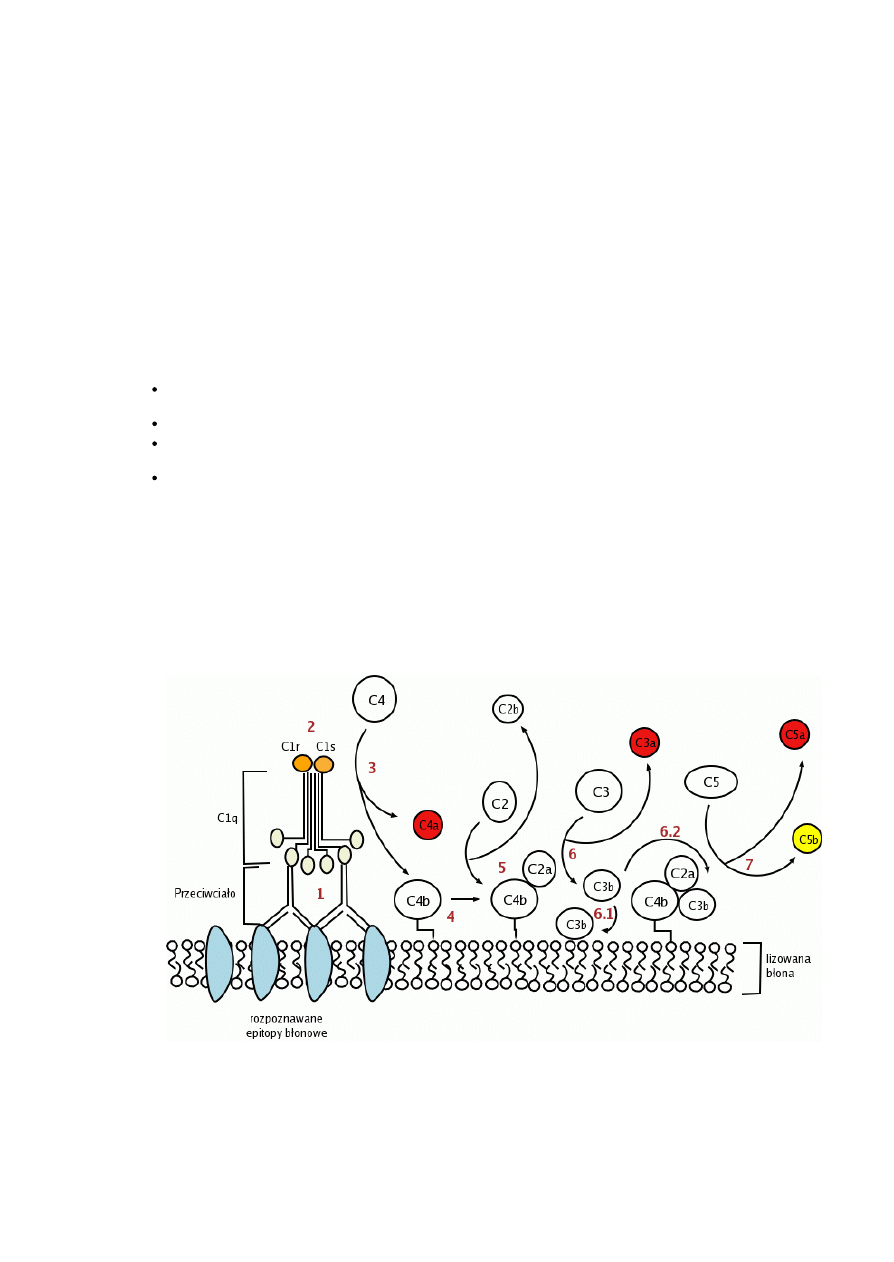
Rys.1. Droga klasyczna aktywacji dopełniacza.
Kolejność zdarzeń w aktywacji dopełniacza drogą klasyczną (kolejne punkty odpowiadają numerom na rysunku):
1. Fragmenty Fab przeciwciał rozpoznają epitopy na powierzchni drobnoustrojów. Cząsteczki C1q dopełniacza przyłączają się do fragmentów Fc
przeciwciał. Do aktywacji dopełniacza konieczne jest połączenie przynajmniej dwóch (z sześciu) domen C1q z dwoma fragmentami Fc.
2. Interakcje przeciwciałaC1q wywołują zmianę konformacyjną cząsteczki C1q, co z kolei pobudza C1r oraz C1s proteazy serynowe. C1r ulega
autokatalizie, następnie przecina, i tym samym uaktywnia, proteazę C1s. Aktywne białko C1s ma zdolność rozkładu czynników C4 i C2.
3. Aktywna proteaza C3 rozkłada C4 do C4a i C4b. C4a jest uwalniane do środowiska reakcji (osocza lub płynu tkankowego).

11.05.2013
Oczos04 - BioInfoArticles
bioinfo.mol.uj.edu.pl/articles/Oczos04
2/4
4. Fragment C4b łączy się z błoną komórkową bakterii.
5. Białko C2 przyłącza się do C4b, w efekcie proteaza C1s rozkłada C2 do C2a i C2b (uwalniany do środowiska reakcji). Powstaje kompleks
C4b2a, nazwany konwertazą C3.
6. Konwertaza C3 rozkłada składnik C3 do C3a (uwalniany do środowiska reakcji) oraz C3b, który może przyłączyć się do błony komórkowej
patogenu i funkcjonować jako opsonina lub przyłączyć się do konwertazy C3, tworząc konwertazę C5.
7. Konwertaza C5 rozkłada białko C5 do C5a (uwalniany do środowiska reakcji) oraz C5b. Ten drugi fragment bierze udział w tworzeniu MAC.
Niezwykle istotne dla prawidłowego działania dopełniacza są konwertazy. Ich znaczenie wynika przede wszystkim z wzmacniającego działania:
pojedyncza konwertaza C3 może potencjalnie wyprodukować setki tysięcy cząsteczek C3b, z których każda może dać początek kolejnej konwertazie C3
lub C5. Z kolei konwertaza C5 może wyprodukować znaczne ilości C5b, a każda z tych cząsteczek może potencjalnie utworzyć nowy MAC. Podobną
rolę odgrywa także proteaza C1s, która może dostarczyć dużych ilości C4b.
Droga alternatywna (properdynowa)
Droga alternatywna rozpoczyna się spontanicznie (bez udziału przeciwciał) i atakuje każdą dostępną błonę biologiczną z wyjątkiem błon komórek
własnych organizmu, na których jest unieszkodliwiana.
Rys.2. Droga alternatywna aktywacji dopełniacza.
Kolejność zdarzeń w aktywacji dopełniacza drogą alternatywną (kolejne punkty odpowiadają numerom na rysunku):
1. W osoczu występuje białko C3(H
2
O), będące pobudzoną formą C3. Ta cząsteczka może wiązać czynnik B, który w obecności jonów magnezu
oraz czynnika D jest rozbijany na fragmenty Ba (uwalniany do środowiska reakcji) oraz Bb
2. Fragment Bb pozostaje związany z C3(H
2
O). Powstały kompleks jest aktywny enzymatycznie i tworzy rozpuszczalną konwertazę C3 drogi
alternatywnej.
3. Rozpuszczalna konwertaza rozbija białko C3 do C3a oraz C3b, który może się przyłączać do błony komórkowej.
4. Związany z błoną C3b przyłącza czynnik B, który przy udziale czynnika D jest rozbijany na Ba i Bb. Fragment Bb pozostaje związany z C3B,
w ten sposób powstaje związana z błoną konwertaza C3 drogi alternatywnej (C3bBb), która jest dodatkowo stabilizowana czynnikiem P, czyli
properdyną. Z tego powodu droga alternatywna nazywana bywa także properdynową.
5. Konwertaza C3 rozkłada białko C3, dając w efekcie C3a i C3b. Czynnik C3b może przyłączyć się do błony. Może również przyłączać się do
kompleksu C3bBb (konwertazy C3), tworząc konwertazę C5 drogi alternatywnej.
6. Konwertaza C5 rozkłada C5 do anafilatoksyny C5a oraz fragmentu C5b, który zapoczątkuje tworzenie MAC.
Droga lektynowa
Droga lektynowa jest w ogólnych zarysach podobna do drogi klasycznej. Różnią się one tylko pierwszymi etapami. W przypadku drogi lektynowej
obecność przeciwciał nie jest potrzebna. Przeciwciała są zastąpione kolektynami, białkami nieswoiście wiążącymi cukry na powierzchni drobnoustrojów.
Do kolektyn należą m.in. białka surfaktantu płucnego A i D oraz lektyna wiążąca mannozę (MBL), występująca w osoczu.
Lektyna wiążąca mannozę jest głównym czynnikiem zapoczątkowującym drogę lektynową. Podobnie jak C1q ma ona 6 domen globularnych
umieszczonych na długim styliku, który może wiązać proteazy serynowe MASP1 i MASP2. Gdy nastąpi związanie MBL do powierzchni antygenu,
MASP1 zostaje aktywowana na skutek zmiany konformacyjnej trzonka MBL. Aktywna MASP1 może dokonać proteolitycznego cięcia MASP2
(analogia do aktywacji proteazy C1s przez C1r). Ten enzym z kolei jest odpowiednikiem C1s i może rozkładać C2 i C4. Dalsze etapy są identyczne jak
w klasycznej drodze aktywacji dopełniacza.
Droga alternatywna i droga lektynowa mają duże znaczenie w walce z patogenami, mogą bowiem zapoczątkować reakcję odpornościową bezpośrednio
po wniknięciu drobnoustroju do organizmu. Podczas gdy droga klasyczna może być rozpoczęta dopiero na skutek wytworzenia przeciwciał, co
następuje po upływie pewnego czasu od pojawienia się antygenu w organizmie.
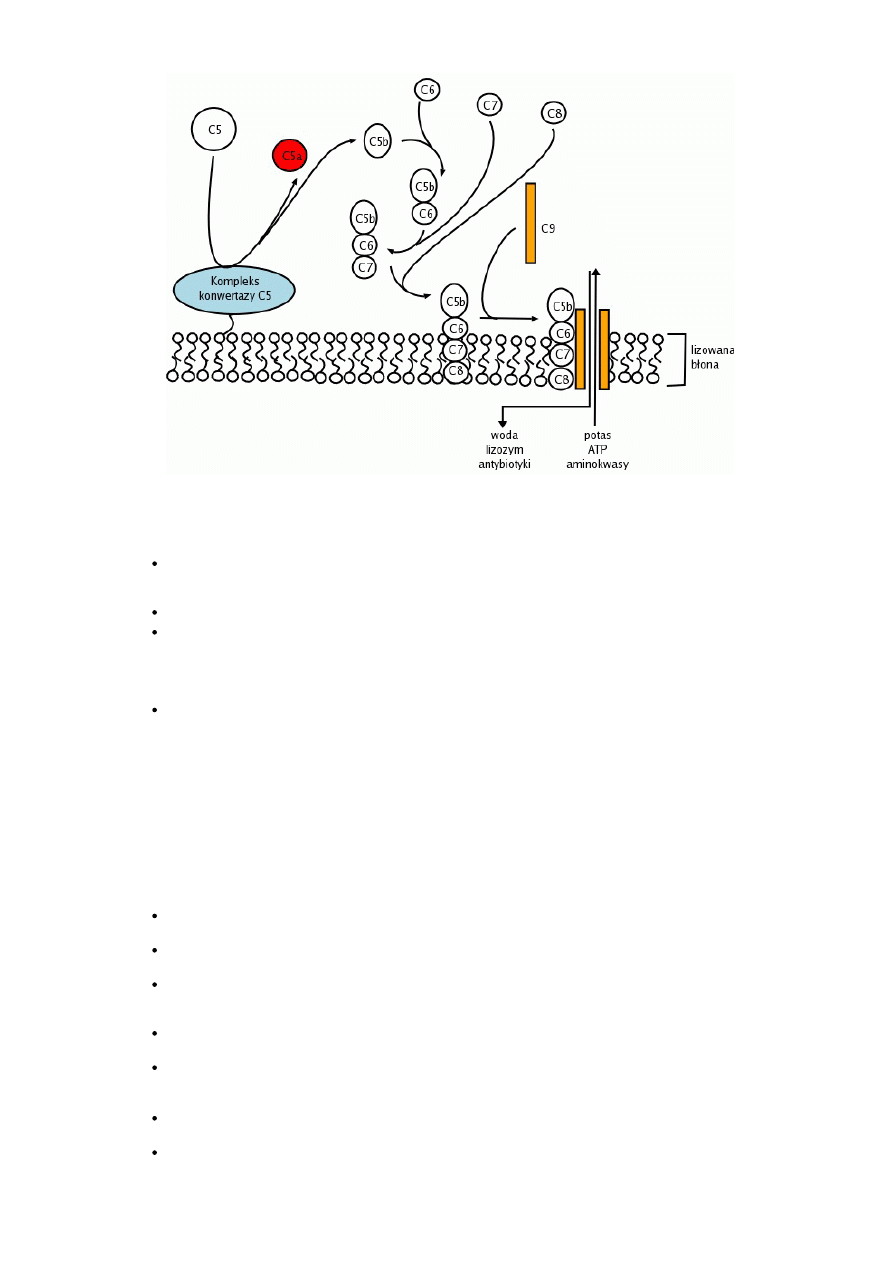
Tworzenie kompleksu atakującego błonę
Tworzenie kompleksu atakującego błonę MAC (membrane attacking complex) składa się wyłącznie z reakcji nie wymagających aktywności
enzymatycznych. Po wytworzeniu C5b przez konwertazę dowolnej z dróg aktywacji dopełniacza, dochodzi do połączenia się C5b z C6. W kolejnym
kroku do C5b6 dołączane są C7 i C8. Powstaje kompleks C5b8, który ma zdolność włączania się w błonę komórkową i przyłączania cząsteczek C9.
Przyłączenie 214 cząsteczek C9 powoduje utworzenie w błonie komórkowej kanału. Powstanie kanałów powoduje wypływ z komórki jonów, ATP,
substancji odżywczych. Równocześnie do komórki napływa woda (ze względu na wyższe ciśnienie osmotyczne w komórce). Mogą się do niej
11.05.2013
Oczos04 - BioInfoArticles
bioinfo.mol.uj.edu.pl/articles/Oczos04
3/4
dostawać także czynniki bakteriobójcze i bakteriostatyczne (np. lizozym) oraz antybiotyki.
Rys.3. Tworzenie kompleksu atakującego błonę.
Funkcje układu dopełniacza
Układ dopełniacza odpowiada za wiele ważnych mechanizmów efektorowych odpowiedzi immunologicznej:
Zapoczątkowanie (ostrego) zapalenia przez bezpośrednią aktywację komórek tucznych (peptydy zapalne C3a i C5a). Wiązanie anafilatoksyn
C3a oraz C5a do receptorów komórek tucznych pobudza degranulację ich ziarnistości i uwalnianie mediatorów farmakologicznych takich jak
histamina, która powoduje skurcz mięśni gładkich i wzrost przepuszczalności naczyń krwionośnych.
Składnik C5a wykazuje aktywność chemotaktyczną, przyciąga neutrofile do miejsca ataku bakterii.
Wzmocnienie wiązania między bakterią i fagocytem ułatwiające pochłonięcie bakterii (opsonizacja). Monocyty, makrofagi, neutrofile i
erytrocyty posiadają na powierzchni receptory dla składników dopełniacza: C3b, C4b. Peptydy C3b oraz C4b pełnią rolę opsonin, opłaszczają
bakterię. Związanie receptorów z opsoninami wzmacnia kontakt bakteriakomórka żerna i ułatwia fagocytozę. Erytrocyty poprzez receptory CR1
(CD35) wiążą kompleksy immunologiczne (ich składnikiem jest C3b), by dostarczyć je do wątroby i śledziony w celu usunięcia przez komórki
żerne.
Zabijanie drobnoustrojów przez kompleks atakujący błonę. Składniki C5b do C9, a w szczególności C9, tworzą pory w błonie komórki
docelowej. Poprzez te kanały transbłonowe następuje wyciek wewnątrzkomórkowych składników i napływ wody do wnętrza komórki. W
rezultacie komórka ulega lizie.
Regulacja aktywności dopełniacza
Dopełniacz ze swoją zdolnością indukowania fagozytozy, degranulacji komórek tucznych z uwolnieniem bardzo aktywnych biologicznie czynników,
zdolnością wywoływania lizy komórki poprzez tworzenie MAC jest groźną bronią i bez mechanizmów regulujących jego aktywność, łatwo mógłby
prowadzić do uszkodzenia naszych własnych komórek.
Mechanizmy i czynniki regulujące aktywność układu dopełniacza można podzielić na takie, które są związane z błonami komórkowymi oraz takie, które
działają w płynach tkankowych, głównie w osoczu.
Czynniki regulujące obecne w błonach komórkowych:
CR1 (CD35) kofaktor dla czynnika I rozkładającego C4b i C3b. Inaktywuje także konwertazy C3 i C5 (czynniki C3b i C4b są ich
składnikami), chroniąc komórki mające na powierzchni CR1 przed "przypadkową lizą".
DAF (CD55) decay accelerating factor wybitnie przyspiesza spontaniczny rozpad konwertaz C3 i C5 obydwu dróg. Działa przeciwnie do
properdyny. Występuje na krwinkach, komórkach śródbłonka naczyń i wielu innych komórkach.
MCP (CD46) membrane cofactor protein wiąże składniki C3b i C4b będące w stanie wolnym lub obecne w konwertazie. Jest kofaktorem
dla czynnika I. Podobnie jak DAF zapobiega formowaniu na powierzchni błony konwertaz C3. MCP jest obecny prawie na wszystkich
komórkach jądrzastych.
HRF (C8bp C8 binding protein) hamuje polimeryzację C9, formowanie się kompleksu atakującego błonę MAC i tworzenie się kanałów.
HRF występuje na limfocytach T i B, neutrofilach i monocytach.
Protektyna (CD59) analog HRF, występujący między innymi w błonie komórkowej wszystkich krwinek i plemników.
Czynniki osoczowe regulujące aktywność dopełniacza:
Inhibitor C1 wiąże aktywny C1r i C1s blokując dalszą aktywację. Zabezpiecza organizm przed konsekwencjami spontanicznej aktywacji. Silne
aktywatory klasycznej drogi aktywacji dopełniacza (np. kompleksy immunologiczne) przełamują tę blokadę.
Czynnik I inaktywator C3b/C4b jest proteazą osocza rozkładającą C3b i C4b zarówno wolne jak i związane w konwertazach. Do aktywności
wymaga kofaktorów. Kofaktorem czynnika I w błonach są CR1, CR2 oraz MCP, a w płynach tkankowych czynnik H oraz białko wiążące C4.

11.05.2013
Oczos04 - BioInfoArticles
bioinfo.mol.uj.edu.pl/articles/Oczos04
4/4
Articles: Oczos04 (last edited 20110215 23:03:59 by
Białko wiążące C4 wiążąc C4b, przyspiesza rozpad konwertazy C3 drogi klasycznej (zarówno rozpad spontaniczny jak również wywołany
przez czynnik I).
Czynnik H wiążąc C3b, przyspiesza rozpad konwertazy C3 drogi alternatywnej.
Witronektyna (białko S) wiąże się z kompleksem C5b7 i uniemożliwia polimeryzację C9. Hamuje łączenie się C5b7 z błoną komórkową.
Literatura
J. Gołąb, M. Jakóbisiak, W. Lasek (red.), "Immunologia", Wydawnictwo Naukowe PWN, Warszawa 2004
P. M. Lydyard, A. Whelan, M. W. Fanger, "Krótkie wykłady: Immunologia", Wydawnictwo Naukowe PWN, Warszawa 2001
J. Brostoff, D. Male, I. Roitt, "Immunologia", Wydawnictwo Lekarskie PZWL, Warszawa 2000
http://microbiology.univ.gda.pl/wyklad3.pdf
http://pl.wikipedia.org/wiki/Dope%C5%82niacz_%28biologia%29
Kategorie: Immunologia
Wyszukiwarka
Podobne podstrony:
Wykład 3, 4 układ dopełniacza
UKŁAD DOPEŁNIACZA
Układ dopełniacza
UKŁAD DOPEŁNIACZA
Uklad pokarmowy
Układ mięśniowy
układ moczowy
Układ nerwowy
oddechowy uklad
Uklad oddechowy2
T5 UKŁAD HYDRAYLICZNY PODNOSZENIA OSPRZĘT DODATKOWY
UKŁAD PŁCIOWY MĘSKI ptt
układ naczyniowy wstep
Uklad oddech wyklad
W 11 Leki działające pobudzająco na ośrodkowy układ
UKŁAD MOCZOWY WYWIADY
więcej podobnych podstron