
4
Behawlor
a przystosowanie
• do otoczenia
Badanie adaptacji zachowania do warunków środowiskowych można
najlepiej prześledzić, obserwując zwierzęta żyjąccw warunkach ekstremalnych
- na pustyniach i w rejonie arktycznym. W obu przypadkach podstawową
trudność dla zwierzęcia stanowią warunki termiczne, związane nie tyle z mak
symalną temperaturą otoczenia, ile z jej okresowymi i dobowymi wahaniami.
W rejonach arktycznych np. zmiany temperatury w ciągu roku osiągają 20-60°C,
natomiast na pustyniach w ciągu doby temperatura może wahać się o l(Mł5°C
(Hardy, 1972).
W warunkach tych nieco inne kłopoty mają zwierzęta zmiennocieplne
i stałocieplne. Generalnie jednak można powiedzieć, że w środowisku pustyn
nym chodzi z jednej strony o zabezpieczenie się przed przegrzaniem i odwod
nieniem, z drugiej (zwłaszcza u gatunków zmiennocieplnych) - o zachowanie
odpowiedniej ilości ciepła w czasie chłodnej nocy. Na terenach arktycznych
zwierzęta muszą uchronić się przed oziębieniem i zamarznięciem. W obydwóch
strefach dużym problemem może być zdobycie pokarmu.
W warunkach ekstremalnych podstawową strategią behawioralną jest odpo
wiednie dostosowanie rytmu aktywności do zmian zachodzących w otoczeniu
(Hamilton, 1973). Wiąże się to z zasygnalizowaną w rozdziale 2 ogólnie regu
lacyjną funkcją behawioru. Zwierzęta unikają najdrastyczniejszych warunków
termicznych, chroniąc się przed Słońcem w cieniu lub przed chłodem w norach
i innych kryjówkach. U pustynnych zwierząt nomadycznych mogą nastąpić
czasowe przerwy w aktywności, wymuszone groźbą przegrzania. Na przykład,
dla szarańczy dłuższy lot w temperaturze 38°C jest niemożliwy ze względu
na szybkie zwiększanie się ciepłoty ciała w czasie aktywności do letalnego
poziomu 45°C. Dlatego też owady te co jakiś czas wypoczywają i przywracają
temperaturę ciała do normalnego poziomu (Hardy, 1972).
Nieco inny typ przystosowania behawioralnego wykazują saharyjskie
mrówki pustynne z rodzaju Cataglyphis. Są one wśród owadów prawdziwymi
championami w znoszeniu wysokiej temperatury - potrafią poruszać się na
powierzchni o temperaturze dochodzącej do 70°C, a ich ciała ogrzewają się do

91
poziomu przekraczającego 50°C. Mrówki te poruszają się jednak z ogromną
prędkością (1 m/sek.), co ogranicza ekspozycję owadów na działanie Słońca,
a jednocześnie ułatwia oddawanie ciepła przez konwekcję (Wehner i in., 1992).
Podobną strategię stosuje suseł pustynny - Ammospermophilus leucurus, który
dosłownie przebiega między „strefami cienia", po drodze pospiesznie żerując
i rozglądając się za ewentualnym drapieżnikiem (Chappell i Barholomew, 1981).
Charakter przystosowania behawioralnego ma również wiele migracji sezo
nowych. U ptaków przebiegają one między północnymi a południowymi kon
tynentami globu. Zwierzęta unikają tym samym zimy na północy, nie chodzi
tu jednak głównie o warunki termiczne, lecz raczej o drastyczne niedobory
pokarmu, które występują w tym czasie (Bright, 1993). U ptaków występują
także migracje krótsze (w ramach jednej strefy klimatycznej), które wynikają
z lokalnego braku pokarmu lub wody. Dzięki możliwości poruszania się na
większych dystansach ptaki radzą sobie nieźle w środowisku pustynnym
(Pough i in., 2002).
Krańcowo odmiennym sposobem przystosowania się do ekstremalnych
warunków i związanych z nimi niedoborami pokarmu jest przejście tego trud
nego okresu w stanie uśpienia. Niektóre drobne ssaki z grupy owadożernych,
gryzoni i nietoperzy, żyjące na półkuli północnej, zawsze w tym samym okresie
-jesienią wchodzą w stan hibernacji właściwej lub obligatoryjnej. Inne reagują
w ten sposób nieregularnie na niedobory pokarmu i na spadek temperatury
poniżej pewnego punktu. Jest to hibernacja fakultatywna. Duże zwierzęta dra
pieżne, jak np. niedźwiedzie czy borsuki, zapadają w zimowy letarg, natomiast
uzimujących gadów obserwuje się odrętwienie (Lawrence, 1995).
Hibernacja właściwa poprzedzana jest okresem intensywnego żerowania
dla zgromadzenia odpowiedniej ilości substancji zapasowych w postaci spe
cjalnego tłuszczu. Przebiega ona w kryjówce dobrze izolowanej od otoczenia.
U niektórych gatunków zapadanie w sen trwa kilka dni poprzez serię „próbnych
drzemek", u innych - w ciągu kilku godzin. Hibernacja często nie ma charak
teru ciągłego, lecz jest „rwana". Zwierzęta budzą się kilka razy na pewien czas
(w skali od godzin do dni). Wówczas też następuje wydalanie, a czasem zjada
nie zgromadzonego w norze pokarmu (Hardy, 1972).
W czasie hibernacji właściwej do wielkości minimalnej (1/70) maleje tempo
przemiany materii, liczba oddechów i uderzeń serca, temperatura ciała jest tylko
nieznacznie wyższa od temperatury otoczenia. Wyhamowaniu ulega także
funkcjonowanie niektórych gruczołów wydzielania wewnętrznego. Jednak hiber
nujące zwierzęta zaopatrzone są w fizjologiczny mechanizm kontrolny. Jeśli
temperatura otoczenia niebezpiecznie spada, zostaje zaktywizowane tempo
przemiany materii albo zwierzęta te budzą się (Kayser, 1961).

92
Odrętwienie u gadów różni się od właściwej hibernacji brakiem owego
mechanizmu kontrolnego. W procesie zasypiania i budzenia się zwierzę jest
pasywne i ściśle uzależnione od wahań temperatury. Jednak gady, podobnie jak
ssaki, poszukują do zimowania nor, jam, wykrotów itd. (często zresztą wielu
przedstawicieli jednego gatunku gromadzi się w tych miejscach). Faza odrę
twienia u zwierząt zmiennocieplnych może być bardzo długa. U północnoame
rykańskiego węża Thamnophis sirtalis w Kanadzie przeciąga się ona na niemal
osiem miesięcy - od października do połowy maja. Ma doniosłe konsekwencje
dla biologii tego gatunku, a zwłaszcza dla jego cyklu rozrodczego. Kopulacja
odbywa się zaraz po wyłonieniu się wężów z nor, a wówczas poziom hormo
nów płciowych zarówno samczych, jak i samiczych jest niski. Także gamety
tworzone przez osobniki obu płci nie są należycie rozwinięte. Aktywność
płciowa u Thamnophis sirtalis jest więc rozkojarzona z cyklem hormonalnym
(Crews, 1987).
Odpowiednikiem hibernacji jest estywacja, która przebiega w warunkach
wysokiej temperatury i suszy. Dla wielu zwierząt (niektóre mięczaki, ryby i ssaki)
pokarm jest wówczas niemożliwy do zdobycia i przespanie tego trudnego okresu
staje się praktycznie jedynym sposobem przetrwania.
Oprócz termoregulacji behawioralnej związanej z aktywnością spotykamy
jeszcze inne rodzaje zachowań, mające znaczenie przystosowawcze w wyżej
omawianych warunkach środowiskowych (Hamilton, 1973; Gautier-Pithers
i Dagg, 1981; Kaleta, 1998; Pough i in., 2002):
1. Powiększanie i zmniejszanie powierzchni ciała
Dodatkowe powierzchnie pozwalają w razie konieczności szybciej pozbyć
się nadmiaru ciepła albo wręcz przeciwnie - ciepło to pozyskać. W klimacie
gorącym dobrym przykładem takich dodatkowych powierzchni są uszy słonia
afrykańskiego (którymi może się wachlować).
W klimacie chłodnym ograniczanie powierzchni widoczne jest np. u lisa
polarnego (Alopex lagopus) w czasie snu. Jego zwinięte w kłębek ciało przy
pomina kulę, a w dodatku przykrywa się - niczym kołdrą - gęsto owłosionym
ogonem.
2. Eksponowanie ciała
Obserwowane jest ono zarówno u zwierząt zmiennocieplnych (np. jasz
czurki), jak i stałocieplnych (naczelne). Wiąże się z przyjęciem takiej pozycji
ciała, przy której jak najszybciej się ogrzewa. U małp jest to na przykład ekspo
nowanie w kierunku Słońca części ciała nieowłosionych lub słabiej owłosionych,
jak choćby dłoni.

93
R y s u n e k 8. Jedna z behawioralnych strategii ochrony przed operacją słoneczną - osłanianie się
ogonem u wiewiórki ziemnej kapskiej (za Pough i in., 2002)
3. Użycie osłony
Wiewiórka ziemna kapska (Xenus inauris) niczym parasolem osłania ciało
przed operacją słoneczną (rys. 8).
4. Zmiana barwy
Jasna barwa powierzchni ciała powoduje odbijanie promieni słonecznych,
natomiast ciemna - pochłanianie ich. Strategia zmiany barw obserwowana jest
u zwierząt zmiennocieplnych. Jeden z jej mechanizmów to obserwowana u nie-

94
których jaszczurek zmiana ciemniejszej barwy, występującej rano, na jaśniejszą
w południe.
5. Zmiana położenia sierści/piór
Gdy sierść/pióra są położone, pełnią funkcję izolacyjną, natomiast gdy są
nastroszone - ochładzają. Ten typ termoregulacji behawioralnej jest doskonale
widoczny u kondorów i sępów, które w zależności od konieczności stroszą lub
kładą kryzę składającą się z długich piór, okalających szyję.
6. Ziajanie
U psa domowego, nie posiadającego w zasadzie gruczołów potowych, wystę
puje zachowanie, które umożliwia oddawanie ciepła z jamy śluzowej otwartego
pyska przy jednoczesnym wykonywaniu przez zwierzę szybkich oddechów.
Analogiczne zjawisko występuje u żyjących na półpustyniach i pustyniach
antylop, z tym że „ziajanie" odbywa się poprzez nozdrza (Estes, 1991).
7. Taplanie się w błocie
Dla dużych ssaków, o powierzchni ciała pokrytej rzadkim włosem (świnio-
wate, hipopotamy, nosorożce), kąpiele błotne są znakomitym środkiem wspo
magającym termoregulację. Na ich ciele odkłada się bowiem warstwa izolacyjna
z przyschniętego błota.
8. Ograniczone wydalanie moczu
Mocz wydalany jest rzadziej, w postaci zagęszczonej. Zjawisko obserwowane
jest u wielu gatunków zarówno kopytnych, jak i gryzoni, żyjących na półpusty
niach i pustyniach.
9. Termoregulacja socjalna
Tulenie się do siebie w czasie hibernacji obserwowane np. u niektórych
gryzoni sprzyja zahamowaniu utraty ciepła. Wielbłądy {Camelus dromedarius)
też wypoczywając, leżą tuż obok siebie, ale tym razem chodzi o ograniczenie
dostępu ciepła słonecznego do powierzchni ciała.
10. Pobieranie pokarmu
Żyjące na pustyniach ssaki kopytne zadowalają się praktycznie wszystkimi
rodzajami roślin. W związku z tym np. u wielbłądów (rodzaj Camelus) uformo
wały się ewolucyjnie grube wargi, które umożliwiają mu bezpieczne ogryzanie
roślin o szorstkich, włoskowatych liściach. Lis polarny żyjący na arktycznej tun
drze też wykorzystuje wszystkie możliwe rodzaje pokarmu zwierzęcego i roślin
nego, chętnie zjada padlinę, a w skrajnych przypadkach nawet własne odchody.

95
Liczne i interesujące przykłady przystosowania do otoczenia o charakterze
zachowań można zaobserwować u zwierząt morskich. Woda morska jest specy
ficznym środowiskiem, którego czynniki w sposób niekorzystny mogą wpływać
na przeżycie organizmów. Oto niektóre z tych parametrów (Levinton, 1995):
1. Temperatura
Znakomita większość zwierząt morskich to organizmy zmiennocieplne,
których temperatura ciała uzależniona jest od wody. U wielu z nich temperatura
ta wpływa na rozród, na zmianę reprodukcji z aseksualnej w seksualną, a nawet
determinuje płeć. Sezonowe zmiany temperatury wody w ciągu roku mogą
zmusić zwierzęta do zahamowania rozrodu w cieplejszych miesiącach wiosen
nych i letnich, jak w przypadku kalifornijskiej rozgwiazdy - Pisaster ochraceus.
2.
Zasolenie
Organizmy morskie muszą aktywnie regulować zawartość soli w płynach
ustrojowych. Jest w nich mniejsze stężenie soli aniżeli w otaczającej wodzie,
toteż aby zapobiec utracie płynów, ryby połykają wodę, usuwając jednocześnie
nadmiar soli za pomocą skrzeli.
3. Tlen
Warunki tlenowe w rozmaitych miejscach akwenu morskiego są różne
(np. lepsze na powierzchni niż w głębinach). Zwierzęta poruszające się mogą
po prostu unikać miejsc słabiej natlenionych; jeśli zmuszone są w nich przeby
wać, ograniczają aktywność.
4. Światło
Zwierzęta żyjące bliżej powierzchni wykorzystują światło słoneczne
w orientacji. Te przebywające w głębinach tworzą własne światło dzięki zjawisku
bioluminescencji,
które zachodzi dzięki działaniom symbiotycznym wobec ryb,
bakterii lub wyspecjalizowanych struktur ciała. Bioluminescencja jest bardzo
rozpowszechniona w świecie zwierząt morskich i występuje praktycznie we
wszystkich grupach systematycznych - począwszy od pierwotniaków, a skoń
czywszy na rybach. Jej funkcja u zwierząt morskich nie jest do końca wyja
śniona. Przypuszczalnie służy przede wszystkim do dezorientowania drapież
ników, przywabiania zdobyczy, kamuflażu i komunikacji (Levinton, 1995;
Burchett, 1996).
5. Pływy morza
Ruch fal może wymusić aktywność niektórych organizmów. Odpływ powo
duje np., że wiele skorupiaków wycofuje się w tym czasie do swoich zachowu
jących wilgoć nor, by uniknąć wyschnięcia ciała.

96
Na koniec warto dodać, że u ssaków morskich występuje zdumiewające
zachowanie: długotrwałe nurkowanie połączone z zatrzymaniem oddechu. Rekor
dzistami są: wśród płetwonogich - słoń morski południowy (Mirounga konina),
który nurkuje na głębokość około 1500 m i trwa to dwie godziny; wśród waleni
- kaszalot, u którego te dwa wskaźniki wynoszą odpowiednio 3000 m i 138
minut. Zjawisku towarzyszy wiele zmian anatomicznych i fizjologicznych
(zapadanie się płuc, bradykardia, zapadanie w sen itd.). Zachowanie to obser
wuje się głównie w czasie żerowania. Jednak istnieje teoria, że częste, głębokie
nurkowanie u słonia morskiego może służyć oszczędzaniu energii przez to
zwierzę. Badania potwierdziły bowiem, że koszt energetyczny nurkowania jest
w tym przypadku mniejszy, niż nawet bierne unoszenie się na powierzchni
(Berta i Sumich, 1999).

5.
Pomyłki i trudności
w badaniu zachowania
się zwierząt
Jak w każdej dyscyplinie badawczej, tak i w badaniu zachowania się zwie
rząt oprócz sukcesów odnotowano także spektakularne pomyłki. Powody ich są
oczywiście różnego typu, jednak błędy we wnioskowaniu i brak należytej
ostrożności w uogólnianiu wyników obserwacji należą chyba do najczęstszych.
Do tego dochodzą problemy związane z techniką badawczą (np. sposób obser
wacji i rejestrowania ich wyników, mierzalność cech jakościowych behawioru
itd.) oraz trudności interpretacyjne. Te ostatnie wynikają z ludzkiej skłonności
do antropomorfizacji (tłumaczenie zachowania się zwierzęcia analogicznie do
ludzkiego) bądź z potrzeby wyjaśniania mechanicystycznego (traktowanie
zachowania się zwierzęcia na podobieństwo pracującej maszyny jako zbioru
zautomatyzowanych reakcji). •
Poniżej zostaną zaprezentowane jedynie wybrane przykłady sytuacji, w któ
rych doszło do mniej lub bardziej spektakularnych pomyłek w ocenie zachowa
nia się zwierząt.
Jak się wydaje, jedną z charakterystycznych cech błędnego wnioskowania
w interpretacji behawioru zwierząt jest poszukiwanie jakichś wyspekulowanych
wyjaśnień przy jednoczesnym niedostrzeganiu przyczyny najprostszej. Tak np.
na początku XX wieku środowisko naukowe pasjonowało się przypadkami
„mądrych zwierząt", które potrafiły się komunikować z człowiekiem i rozwią
zywać nawet skomplikowane zadania matematyczne. Najsłynniejszym spośród
tych zwierząt był koń, Mądry Hans. Jego właściciel na licznych pokazach pre
zentował niezwykłe umiejętności swojego pupila, który wystukiwał kopytem
liczbę stanowiącą rozwiązanie zadania, posługując się kodem liczbowo-litero-
wym tworzył słowa, a nawet zdania, a także odpowiadał na pytania, kiwając
potakująco albo kręcąc przecząco głową (Katz, 1951). Jednak w trakcie uważ
nych badań, zdolności matematyczne i lingwistyczne Mądrego Hansa zostały
zakwestionowane, gdy okazało się, że koń reaguje po prostu na sygnały nada
wane nieświadomie przez swojego opiekuna. Przy wystukiwaniu kopytem
zwierzę reagowało po prostu na mimowolną zmianę pozycji ciała czy inny,

98
trudny do zauważenia dla człowieka sygnał J ę z y k a ciała" swojego opiekuna
i wtedy przerywało stukanie.
Kiedy rozpoczęły się na szeroką skalę badania terenowe zachowania się
dużych ssaków, rychło okazało się, że aby właściwie rozszyfrować liczne zagadki
związane z obyczajami przedstawicieli poszczególnych gatunków, niezbędne są
długotrwałe studia. Natomiast zadania wyrywkowe, w dodatku niezbyt poprawne
metodycznie, mogą prowadzić do fałszywych wniosków. Tak np. badający
w latach 50. życie społeczne pawiana sawannowego (Papio cynocephalus)
Washburn i De Vore uzyskali rezultat znacząco różny aniżeli ich następcy z lat
70. XX wieku, tacy jak np. S. Strum (Moss, 1982). W pierwszej wersji społe
czeństwo pawianów miało być grupą, w której najważniejszą rolę odgrywają
dojrzałe samce, tworzące rodzaj koalicji, natomiast hierarchia wśród samic
miała charakter niestabilny. Jednak długofalowe obserwacje, prowadzone
przede wszystkim przez badaczkę Shirley Strum, która przebywając z małpami,
na długo dosłownie „wtopiła się" w stado pawianów, zmieniły ten obraz. Okazał
on się znacznie bardziej skomplikowany: zasadniczą rolę w pawianiej społecz
ności odgrywają grupy rodzinne, koncentrujące się wokół dojrzałych samic
i tworzące mniejsze lub większe „strefy wpływów". W tej wersji samce odgry
wają znacznie mniejszą rolę, starają się one zdobyć względy co bardziej wpły
wowych samic.
Tak więc, choć Washburn i De Vore poczynili wiele interesujących obser
wacji dotyczących obyczajów pawiana sawannowego, jednak uzyskany przez
tych badaczy całościowy obraz społeczności tego gatunku okazał się dalece
niekompletny. Nie wynikało to rzecz jasna z jakichś celowych zaniedbań wymie
nionych badaczy, ale z innego podejścia metodycznego do prowadzonych
obserwacji (Moss, 1982).
W badaniu zachowania się zwierząt często staje się przed koniecznością
testowania kilku równorzędnych hipotez, wyjaśniających taki lub inny rodzaj
behawioru. Może się wówczas zdarzyć, że wybór właściwej interpretacji
napotyka na trudności. Tak np. Schmidt (1995) zajmowała się wyjaśnianiem
funkcji zachowania polegającego na rozpościeraniu skrzydeł u kormorana
rogatego (Phalacrocorax aurituś). Ptak ten, siedząc na gałęzi, często rozpościera
skrzydła (całkowicie lub częściowo) i porusza nimi.
Proponowano następujące hipotezy wyjaśniające ów behawior: (1) suszenie
skrzydeł; (2) termoregulacja; (3) ułatwianie procesu syntezy witaminy D;
(4) pielęgnacja piór w czasie pierzenia; (5) ułatwianie pozbywania się pasoży
tów; (6) ułatwianie zachowania równowagi; (7) forma zachowania społecznego
- imponowanie; (8) manifestowanie wobec innych osobników sukcesu łowiec
kiego; (9) pielęgnowanie piór; (10) ocienianie gniazda; (11) przywracanie pió-

99
rom właściwego położenia. Spośród tych wstępnych hipotez dotąd największe
poparcie wśród badaczy analizujących behawior zdobyły (1) i (2). Jednak po
starannej analizie danych obserwacyjnych, dotyczących czasu spędzanego
przez ptaki w wodzie i na gałęzi, L. Schmidt doszła do wniosku, że suszenie
piór nie jest nadrzędnym celem rozpościerania skrzydeł u tego gatunku kormo
rana. Aby więc odpowiedzieć na pytanie o funkcję tego zachowania się, należa
łoby więc starannie przetestować obserwacyjnie kolejne hipotezy, biorąc za
przedmiot badań ptaki żyjące w możliwie różnych populacjach.
Oceniając, jak zmieniały się poglądy badaczy na zachowanie się poszczegól
nych gatunków i tym samym na pełnione przez nie role w naturalnym środowi
sku, nie sposób oprzeć się refleksji, że najdramatyczniejsza zmiana dokonała
się w przypadku ssaków drapieżnych. Hiena cętkowana, uznawania dawniej za
„tchórzliwego padlinożercę", okazała się w istocie jednym z najsprawniejszych
drapieżników sawanny afrykańskiej, podczas gdy jakby na przekór musiano
przyjąć do wiadomości, że padlinożercą chętnie bywa lew („król zwierząt").
Także obraz zachowania się wilka, spośród zwierząt najbardziej chyba
zmitologizowany w oczach Europejczyków, ulegał stopniowej rewizji od
momentu, gdy zwierzę to poddano naukowym badaniom. Polowania wilków na
zwierzynę płową, które dawniej uważano za destrukcyjne dla tej ostatniej,
zyskały nową ocenę, gdy udowodniono, że drapieżniki te selekcjonują przyszłą
ofiarę, często wybierając na zdobycz osobniki chore, słabe czy stare. Łowy
„krwiożerczego" wilka można zatem uznać paradoksalnie za dobroczynne dla
populacji jego ofiary i dla środowiska w ogóle.

6
Kształtowanie
zachowania przez geny
i środowisko
6.1. Problem genotyp - środowisko
Od momentu, gdy odkryto wpływ materiału genetycznego na ujawnianie
się cech biologicznych, a także kiedy została sformułowana teoria doboru natu
ralnego, pojawiła się w nauce tendencja, by wszelkie różnice między- i wewnątrz-
gatunkowe analizować według dwóch przeciwstawnych sobie czynników
sprawczych - genów i środowiska. Określoną cechę widziano jako pozostającą
pod kontrolą genetyczną albo rozwijającą się pod wpływem środowiska. Dało
to początek znanej w historii psychologii kontrowersji: „instynkt czy uczenie
się?" Taka wizja kształtowania różnic - przede wszystkim między ludźmi -
okazała się zresztą atrakcyjna także dla niebiologów. W naukach humanistycz
nych pojawiło się przeciwstawienie tego, co „wrodzone" i co „wyuczone",
„odziedziczone" i „nabyte", „natury" i „kultury".
Dwubiegunowe widzenie istoty człowieka fatalnie zaowocowało w XX
wieku dwiema najbardziej zbrodniczymi w dziejach doktrynami: nazizmem,
który wyolbrzymiał wpływ elementu wrodzonego (teoria rasy), i komunizmem,
którego dokryna obstawała przy wariancie środowiskowym (rola „kolektywu"
w życiu człowieka, „wychowanie przez pracę" itd.).
W widzeniu przyrodników taki obraz opozycji sfery odziedziczalnej i środo
wiskowej został zakwestionowany dzięki rozwojowi genetyki, a zwłaszcza bada
niom nad molekularnymi mechanizmami dziedziczenia. Dziś już nie ulega wąt
pliwości, że w kształtowaniu określonej cechy wpływy genów i środowiska
przenikają się wzajemnie i uzupełniają. Można mówić raczej o komplementarnym
charakterze obu sfer, nie zaś o ich opozycji. Wpływają na to zarówno pewne
specyficzne cechy dziedziczenia na poziomie molekularnym, jak i na wyższym.
Dwie wymienione poniżej cechy można uznać za najistotniejsze:
• geny nie działają w próżni. Pomiędzy syntezą białka a środowiskiem za
chodzą interakcje poprzez układ neurohormonalny. Procesy te są waż
ne dla rozwoju (ontogenezy) organizmu i dla utrzymania jego funkcji;

101
• wpływ środowiska na cechę może zależeć od podłoża genetycznego
(np. istnieć mogą predyspozycje do uczenia się).
Różne genotypy dają często odmienne efekty w tym samym środowisku,
ale też ten sam genotyp może zaowocować rozmaitymi rezultatami w dwóch
różnych środowiskach. Tak więc ścisłe rozgraniczanie wpływu sfery genetycz
nej i środowiskowej wydaje się być bezzasadne. Biorąc to pod uwagę, w niniej
szym rozdziale procesy dziedziczenia zachowania się i jego „dojrzewania"
osobniczego (przebiegające pod znaczącym wpływem środowiska) zostaną
omówione w jednym rozdziale.
6.2. Genetyka behawioru i relacja
geny - zachowanie się
Komplementarność wpływu dziedziczenia i środowiska nie oznacza, że
w przypadku poszczególnych cech nie można szacować, na ile podlegają one dzia
łaniu genów. Istnieją procedury doświadczalne i matematyczne pozwalające ten
szacunek określić. Między innymi tym właśnie zajmuje się genetyka behawioru.
Mówiąc o dziedziczeniu zachowania się, trzeba przede wszystkim stwierdzić,
że ma ono pewną specyfikę odróżniającą od schematów typowych dla cech
morfologicznych. Wzorce behawioru są bowiem generalnie znacznie mniej
„sztywne" od cech budowy ciała i bardziej uwrażliwione na wpływ środowiska.
Co więcej, niektóre spośród nich, jak na przykład związane z poszukiwaniem
pokarmu, mogą ulegać zmianie praktycznie przez całe życie zwierzęcia.
Badania wpływu genów na zachowanie się mają także dość istotne ograni
czenia w zakresie techniki eksperymentatorskiej. Po pierwsze - mogą być pro
wadzone praktycznie tylko na niektórych gatunkach zwierząt, a mianowicie: na
charakteryzujących się szybką rotacją międzypokoleniową. Chodzi o to, że
niezbędne przy badaniach genetycznych procedury selekcji wymagają śle
dzenia określonej cechy przez większą liczbę pokoleń, a staje się to trudne
u gatunków, które dojrzewają późno i zaczynają się rozmnażać, powiedzmy, po
kilku latach. Dlatego też większość badań dotyczących genetyki behawioru
prowadzi się na gryzoniach laboratoryjnych i muszce owocowej - Drosophila
melanogaster.
Dodatkowym atutem tych zwierząt jest także fakt, iż technicznie
łatwo jest wywołać u nich mutację, która pozostaje jedną z najważniejszych
metod eksperymentalnego wywołania zmiany zachowania się zwierząt.

102
Po drugie - celem zminimalizowania wpływu środowiska w genetyce
behawioru stosuje się przede wszystkim badania laboratoryjne, przy możliwie
pełnej kontroli warunków otoczenia zwierząt.
Geny nie wywołują behawioru w sposób magiczny. Drogę od genu do
zachowania się wyznacza często skomplikowany szlak, obejmujący zarówno
struktury anatomiczne, jak i procesy biochemiczne i fizjologiczne. Dokładne
określenie mechanizmu oddziaływania genów na behawior jest więc z reguły
trudne. Wiadomo, że na poziomie molekularnym określa je np. kodowanie bia
łek, które wchodzą w skład enzymów i hormonów (mających wpływ na za
chowanie się). Na przykład, brak jednego z enzymów powoduje u ludzi choro
bę fenyloketonurię, która powoduje upośledzenie umysłowe (Dewsbury, 1978).
Ulegające w wyniku mutacji zmianie geny mogą również (na wyższym pozio
mie organizacji) modyfikować strukturę narządów, które są związane
z behawiorem. Na przykład, forma bezskrzydła muszek owocowych nie tylko
nie jest w stanie sprawnie poruszać się, ale także tworzyć wibracje, niezbędne
u samców w czasie zalotów. Podobne zmiany związane z dysfunkcją lub zmianą
zachowania się można wywołać mutacyjnie w mięśniach, narządach zmysłu
czy w centralnym układzie nerwowym (Huntingford, 1984).
Interesującą i płodną metodą identyfikacji relacji gen - behawior okazało się
także wykorzystanie mozaiki. Genetyczna mozaika polega na łączeniu u jednego
osobnika tkanek uwarunkowanych przez różne genotypy. Dzięki temu można
wykryć, jaka struktura ciała bezpośrednio odpowiada za dany behawior. Na
przykład, u myszy łączono tkanki pochodzące ze szczepu zabijającego owady
i nie zabijającego. Stosowano różne kombinacje przeszczepiania tkanek i w efek
cie okazało się, że agresywność łowiecką wykazywała mozaika z opuszką
węchową i pniem mózgu pochodzącym ze szczepu zabijającego owady, co
wskazuje na rolę tych struktur w zachowaniu się (Huntingford, 1984).
6.3. Wpływ genów na zachowanie się
Aby ukazać genetyczne podłoże różnic behawioralnych, stosuje się różno
rodne techniki. Jedną z nich jest krzyżowanie ze sobą spokrewnionych gatunków.
W poniższej tabeli 12 przykładowo zestawiono cechy zachowania się bli
sko spokrewnionych ze sobą gryzoni z rodziny marowatych (Caviidae) oraz
behawior ich mieszańca Fj. Mimo iż częstotliwość występowania tych cech
określona jest nie dość precyzyjnie („powszechnie", „rzadko" itd.), analiza
zestawienia dość wyraźnie sugeruje, że mamy do czynienia przynajmniej

103
z niektórymi wzorcami behawioru mającymi znaczący komponent genetyczny.
Wskazuje na to fakt, że np. u mieszańców F, zachowanie się jest często kom
promisem pomiędzy behawiorem form wyjściowych. Ta technika badawcza ma
jednak wiele niedogodności. Mieszańce Fi są często bezpłodne, co uniemożliwia
normalnie stosowane w takiej sytuacji kojarzenie celem uzyskania pokolenia F
2
oraz krzyżowanie wsteczne. Ponadto, stosując tę metodę, zdecydowanie łatwiej
jest zinterpretować zachowanie się organizmów, które mają mniejszą liczbę
chromosomów (dotyczy to np. owadów).
Tabela 12. Niektóre różnice w zachowaniu się dwóch gatunków marowatych (Caviidae) i ich
mieszańców (za Rood, 1972)
Cecha behawioru
Występowanie behawioru u:
aparea
świnki morskiej
mieszańców Fj
(Cavia aparea)
[Cavia
porcellus)
Wokalizacja:
Pokwikiwanie
czasami
powszechnie
powszechnie
Gwizd
brak
powszechnie
brak
Pisk
rzadko
powszechnie
powszechnie
Ćwierkanie
czasami
brak
rzadko
Z a c h o w a n i e agonisty-
czne:
Agresja samca wobec
w o b e c samców i cza
jedynie wobec samców jedynie w o b e c samców
obcych
sem samic
Agresja samicy w o b e c w o b e c samic i czasem
brak
czasami wobec samic
obcych
samców
Agresja samic w o b e c
powszechnie
brak
rzadko
młodych
Czynna defensywa wo powszechnie
czasami
powszechnie
b e c samców
Hierarchia u samic
stabilna
nieokreślona
niestabilna
Z a c h o w a n i e seksu
alne:
Krycie samic nie
brak
czasami
czasami
będących w rui
Krycie
brak
czasami
rzadko
„ h o m o s e k s u a l n e "
Lizanie odbytu
rzadko
powszechnie
powszechnie
W jaki inny sposób można przekonać się, że obserwowany behawior jest
rzeczywiście pod wpływem genów? Odpowiedzi na to pytanie może udzielić
selekcja. Określona reakcja analizowanej grupy zwierząt na sztuczną selekcję
wskazuje na istnienie komponentu genetycznego. Metodyka doświadczenia
polega w tym wypadku na wyborze z populacji osobników charakteryzujących
się skrajnymi wartościami jakiejś cechy. Powiedzmy, że analizowane jest pod
łoże genetyczne cechy A, natomiast A(+) oznacza jej wartości maksymalne,
104
a A(-) wartości minimalne. W każdym pokoleniu wybiera się i kojarzy w za
mkniętej grupie zarówno osobniki A(+), jak i A(-). Jako trzecią prowadzi się
kojarzenia grupy kontrolnej, w której nie ma selekcji. Efekty takich zabiegów
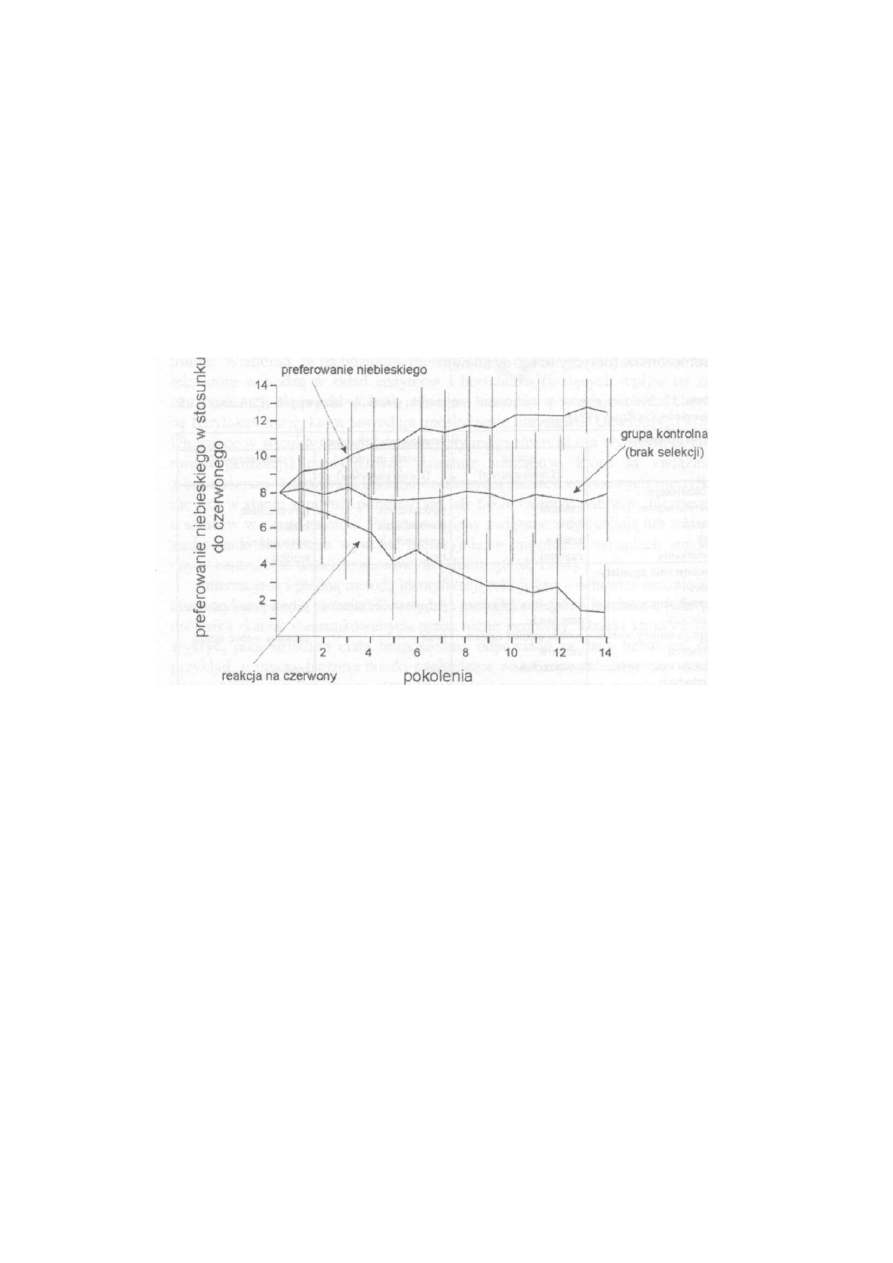
widoczne są na rysunku 9, który przedstawia wyniki selekcjonowania przepió
rek japońskich {Coturnix japonica) na wybraną cechę behawioralną.
R y s u n e k 9. Wyniki selekcji przepiórek na preferencję barw (za Kovach, 1980). Ptaki selekcjonowano
przez 14 pokoleń
Inna metoda polega na tworzeniu jednorodnych genetycznie linii. Opiera
się ona na założeniu, że różnica w behawiorze dwóch wysokozimbredowanych
grup zwierząt, utrzymywanych w jednorodnym środowisku, musi być wynikiem
oddziaływania genotypu. Wysoki stopień zimbredowania uzyskuje się poprzez
kojarzenie w bliskim pokrewieństwie (brat x siostra) i powtarzanie tego przez
przynajmniej 20 pokoleń. Jeśli dwie w ten sposób uzyskane linie wykazują
niezmienne różnice w zachowaniu się, jest oczywiste, że odmienność ta ma
podłoże genetyczne. U myszy laboratoryjnej testowano tą drogą takie zachowa
nia, jak: skłonność do eksploracji, agresywność, zachowanie seksualne, budowa
gniazda (Dewsbury, 1978).

105
W metodzie tej zaistnieć mogą jednak pewne subtelne różnice środowi
skowe, wynikające z wpływu matki na potomka. Aby to zniwelować, można
zastosować technikę odchowu krzyżowego. Polega ona na wychowie potomstwa
przez samice-mamki o odmiennej charakterystyce behawioralnej (np. myszy
agresywne odchowują potomstwo osobników spokojnych i na odwrót). Po
przeprowadzeniu tego zabiegu można dokonać szacunku wpływu mamki na
potomstwo. Jeśli młode nie zmieniło zachowania lub zmieniło je w sposób nie
znaczący, potwierdza to genetyczne uwarunkowanie cechy.
W ślad za samym stwierdzeniem wpływu genotypu na zachowanie się,
w dalszej kolejności pojawia się problem ilości genów zaangażowanych w ten
proces. Wiadomo, że zmianę zachowania się organizmu może wywołać jeden
gen. Wskazują na to wyniki badań prowadzonych między innymi na mutantach
muszki owocowej i pantofelka. W obu wypadkach zmiana behawioru polegała
na pojawieniu się formy patologicznej (np. niewłaściwego sposobu poruszania
się), co wynikało z semiletalnego charakteru mutacji. Podobnie pewne dzie
dziczne choroby ludzkie wywołane są przez jeden zmieniony gen, i im również
towarzyszą zaburzenia zachowań (Grier, 1984). Zmieniony tu gen narusza
skomplikowany proces rozwoju regulowanego faktycznie przez współgranie
wielu genów. Doskonałej metafory, by przedstawić to zjawisko, użył Alcock
(1993). Rolę pojedynczej mutacji porównał on do robotnika pracującego przy
taśmie w fabryce silników samochodowych, który posługuje się niewłaściwym
narzędziem. Dzięki niemu i tylko dzięki niemu końcowy produkt - silnik staje
się wadliwy.
Pojedynczy, zmieniony gen może także działać plejotropowo (wpływać
jednocześnie na różne cechy). Za przykład może służyć tu albinizm. Jak się
okazało, u myszy gen warunkujący tę cechę morfologiczną wpływa zarazem
na wiele złożonych zachowań. Pomysłowe doświadczenie Levine'a (1958)
wykazało np., że samce albinotyczne (wskutek większej agresywności) wygry
wają w konkurencji o samice z rywalami o umaszczeniu dzikim.
Nieczęsto zdarza się badaczom wykryć tak klarowny schemat dziedziczenia
cechy behawioru, jak Rothenbuchlerowi (1964), który badał zachowanie higie
niczne u robotnic pszczół. Behawior ten polega na usuwaniu nieżywych larw
z komórek lęgowych i składa się z dwóch form - otwierania komórki i wycią
gania larwy. Okazało się, że ujawnienie się bądź brak dwóch faz tego zachowa
nia wywołane jest dwoma genami umiejscowionymi w różnych loci i na różnych
chromosomach. Wobec tego istnieją w sumie cztery warianty zachowania:
otwieranie i usuwanie, otwieranie i brak usuwania, usuwanie (w momencie,
gdy komórka zostanie otwarta przez kogoś innego) oraz brak zarówno jednej,
jak i drugiej fazy. Prawidłowe, pełne zachowanie higieniczne robotnicy jest
cechą wywołaną przez geny podwójnie recesywne.

106
Pod kontrolą g e n ó w (nawet pojedynczych) m o ż e być także formuła życia społecznego. U d o w o -
I
dniły to badania na mrówkach przeprowadzone przez badaczy szwajcarskich i amerykańskich. Ich
I przedmiotem była mrówka ognista - Solenopsis invida, gatunek pochodzący z Ameryki Połud
niowej, który „skolonizował" Stany Zjednoczone. Jak się okazało, u tego owada występują dwie
I formy życia społecznego i związane z tym sposoby reprodukcji. W kolonii m o ż e znajdować się
jedna królowa o pokaźnych rozmiarach i pozostawiająca dużą liczbę potomstwa (zjawisko
określane jako monoginia) lub kilka zdecydowanie mniejszych i słabiej rozmnażających się
królowych (poliginia).
Okazuje się, że taki, a nie inny „układ sił" w mrowisku uzależniony jest od genu G
p
-9, którego
I allele oznaczone są literą „b". W rezultacie pod wpływem genotypu BB kształtują się kolonie
monoginiczne, a w wersji Bb - poliginiczne. Badacze ustalili, że mrówki rozpoznają właściwy dla
kolonii rodzaj genotypu. Przy próbie wprowadzenia obcej królowej Bb do mrowiska poliginicznego
jest ona tolerowana, podczas gdy królowa o genotypie BB zostaje po prostu rozerwana na strzępy.
Podobny, smutny los spotyka samicę Bb wprowadzoną do gniazda monoginicznego. Obecnie
trwają spekulacje na temat ewolucji, która stworzyła tak niecodzienną dwoistość struktury
| społecznej u mrówki ognistej (Demarthon, 1998).
Znakomita większość wzorców zachowania się to cechy o rozkładzie cią
głym, które są kształtowane przez wiele genów. Stosowane w tym wypadku
techniki badawcze należą do genetyki cech ilościowych i wykraczają poza ramy
tematyczne niniejszej pracy.
Już dziś pojawiają się poważne sygnały świadczące o tym, że przyszłością
genetyki behawioru zwierząt będą, być może, badania molekularne. Wraz z roz
szyfrowywaniem genomu coraz to nowych organizmów wzrastać będzie także
wiedza o ewentualnym wpływie poszczególnych genów na zachowanie się.
Ponadto już prowadzone badania wskazują także na możliwości genetycznego
modyfikowania behawioru. Na przykład, udało się już doświadczalnie poprawić
uczenie i pamięć myszy poprzez zmianę specyficznego receptora - białka znaj
dującego się w komórkach nerwowych (Tsien, 2000).
6.4. Ontogeneza behawioru
Organizm zwierzęcy rozwija się z połączenia dwóch komórek w złożony
twór składający się z funkcjonalnie zróżnicowanych, choć współgrających ze
sobą tkanek i narządów. W tym obrazie zmian mieści się również rozwój zacho
wania się. W ontogenezie behawior przechodzi zwykle przez kilka faz. U zwie
rząt niższych te fazy rozwojowe manifestują się często w ten sposób, że istnieje
nie tylko odmienność zachowania się, ale również inna morfologia i tryb życia
(np. stadia larwalne u owadów). W przypadku zwierząt wyższych rozwój także

107
nie przebiega jakąś jedną drogą. Na przykład, różne gatunki, by osiągnąć pełnię
dojrzałości, wymagają dłuższego lub krótszego okresu i mniej lub bardziej inten
sywnej opieki rodzicielskiej. W czasie rozwoju uwidacznia się wpływ środowi
ska, które może działać stymulująco, ale i hamująco na różne cechy biologiczne,
w tym również na zachowanie się.
Podstawą rozważań w tym rozdziale będzie ontogeneza behawioru u krę
gowców. Generalnie można podzielić ją na dwie fazy: przed urodzeniem lub
wykluciem (faza prenatalna) i po urodzeniu (faza postnatalna).
W fazie prenatalnej wiele badań przeprowadzono na embrionach kurzych,
u których stwierdzono dość złożoną i systematycznie różnicującą się aktywność
ruchową. Już bardzo wcześnie, bo około 4 dni po rozpoczęciu inkubacji, rozpo
czynają się u embrionu ruchy nieskoordynowane i konwulsyjne, natomiast
bardziej zintegrowana aktywność, prowadząca w końcu do przyjęcia pozycji
wylęgowej, pojawia się około 17. dnia. Stopniowo uaktywniają się też narządy
zmysłów w kolejności, która jest zresztą charakterystyczna dla innych, wyż
szych kręgowców: dotyk, słuch i wzrok. Przed wykluciem pojawia się zdolność
do odbierania bodźców akustycznych z zewnątrz, co ma prawdopodobnie
wpływ na synchronizację lęgu. W momencie wyklucia następuje dramatyczna
zmiana w tonusie mięśni i w miejsce sflaczałego embrionu pojawia się ruchliwe
i żywo reagujące na bodźce pisklę (Dewsbury, 1978).
Już w fazie prenatalnej rozpoczyna się też u zwierząt proces mający zna
mienny wpływ na późniejsze zachowanie się - determinacja płci behawioralnej.
Przez to ostatnie pojęcie rozumie się zachowanie seksualne typowe dla okre
ślonej płci genetycznej. Jak się okazuje, płeć genetyczna i behawioralna nie
muszą być tożsame.
Badania na szczurach wykazały, że kluczowy dla określenia zachowania
seksualnego jest hormon testosteron (Levine, 1966). Jego sekrecjajest znacząca
w okresie 17 dni ciąży do 8-10 dni po urodzeniu, kiedy to następuje prawdopo
dobnie swoiste zorganizowanie mózgu w kierunku zachowania typowego dla
samców (czyli kształtuje się proces maskulinizacji). Oprócz organizacji mózgu
(wpływu na określone jego struktury) testosteron działa po raz drugi, wywołując
aktywizację seksualną samca wobec samicy po osiągnięciu przez zwierzęta
dojrzałości. Kastracja szczura tuż po urodzeniu skutkuje brakiem późniejszej,
właściwej dla samca reakcji wobec samicy. Z kolei wykastrowany samiec, który
jako osobnik dojrzały otrzymuje zastrzyk z hormonu żeńskiego (estrogenu),
wykazuje wobec innego samca lordozę, czyli postawę kopulacyjną typową dla
samicy. Analogicznie do maskulinizacji u samca przebiega feminizacja u samicy;
ona również może ulec zakłóceniu. Zmiany te zostały zestawione w tabeli 13.

108
T a b e l a 13. Procesy kształtowania płci behawioralnej u szczura laboratoryjnego
Oddziaływanie
doświadczalne
po urodzeniu
Oddziaływanie na osobnika dorosłego:
Oddziaływanie
doświadczalne
po urodzeniu
+ testosteron
samiec
+ testosteron
samica
+ estrogen
samiec
+ estrogen
samica
Brak działania
(osobnik
normalny)
właściwy beha-
wior u samca
brak reakcji
brak reakcji
właściwy
behawior u samicy
Kastracja
u s a m c ó w
brak reakcji kopu
lacji w kontakcie
z samicą
pozycja lordozy
w o b e c innego
samca
Podanie
testosteronu
u samic
typowe zachowa
nie seksualne sam
cze (wspinanie się,
intomisja itd.)
brak reakcji lordo
zy w obecności
samca
Można więc powiedzieć, iż determinacja płci behawioralnej polega na
hormonalnej aktywizacji jednego z dwóch ośrodków zawiadujących typowe dla
płci zachowania seksualne.
Pod względem rozwoju w fazie postnatalnej ssaki i ptaki dzielą się na dwie
grupy: gniazdowniki i zagniazdowniki (zob. rozdz. 3). Z punktu widzenia roz
woju zachowania się na uwagę zasługuje przede wszystkim ta druga grupa.
U zagniazdowników proces kształtowania zachowania musi być przynaj
mniej częściowo przyspieszony, choćby dlatego, że młode zwierzę musi często
samodzielnie zacząć pobierać pokarm. Do takich gatunków należy właśnie kura
domowa. Pisklęta kurze zjadają pokarm nawet wtedy, gdy są odizolowane od
kwoki.
Już dość dawno temu ukazano doświadczalnie, jak przebiega ten proces
(Cruze, 1935). Kurczęta mają genetycznie zakodowane zachowanie polegające
na dziobaniu obiektów kontrastujących z podłożem, po którym pisklęta poru
szają się. Początkowo ich uderzenia dziobami są często chybione, jednak aktyw
ność ta ulega doskonaleniu. W doświadczeniu jednej z grup kurcząt umożliwiano
dziobanie podłoża, a innym nie (trzymano je określony czas w ciemności
i żywiono z ręki). Okazało się, że u trzymanych w ciemności młodych ptaków
stopniowo rozwijała się zdolność do bardziej precyzyjnego dziobania. Jednak
proces ten przebiega szybciej wtedy, gdy kurczęta mają możliwość trenowania
celności uderzeń.
Doświadczenie to wskazuje na fundamentalne znaczenie dwóch procesów
zachodzących w fazie postnatalnej: dojrzewania (ang. maturation) i praktyki
(ang. practice). Dojrzewanie polega na osiąganiu pełnej aktywności przez
struktury wpływające na zachowanie się (np. sekrecja hormonów przez gruczoły,

109
zmiany w siatkówce itd.). Praktyka natomiast jest doskonaleniem behawioru
poprzez powtarzanie, czynieniem go w pełni zorganizowanym i kompletnym.
Podstawową metodą badawczą, pokazującą, jak rozwija się behawior
w okresie postnatalnym, są eksperymenty izolacyjne (tzw. deprywacje). Pole
gają one albo na sztucznym odchowie pojedynczych osobników w całkowitej
izolacji od normalnego środowiska, albo na izolacji zwierząt w określonym
czasie od specyficznych bodźców będących impulsem do poszczególnych
zachowań.
Przeprowadzane na różnych gatunkach kręgowców obserwacje pojedyn
czych, izolowanych osobników (gdzie wyeliminowane jest ewentualne naśla
downictwo) pokazują, że ich zachowania pojawiają się stopniowo, w czasowo
określonych fazach. Tak np. behawior lokomotoryczny szczura laboratoryjnego
obejmuje dziesięć podstawowych sprawności i rozwija się w ciągu 22 dni.
Najwcześniejszy wzorzec behawioru pojawia się w pierwszym dniu po urodzeniu
- jest to przyjmowanie pozycji wyprostowanej. Najpóźniejsza aktywność to
schodzenie po linie głową w dół i skoki przez poziome przeszkody (Altman
i Sudarshan, 1975).
U gatunków żyjących społecznie izolacja może wywołać szczególny efekt.
Normalny rozwój u tych zwierząt musi zakładać socjalizację, czyli opanowanie
zachowań pozwalających funkcjonować w grupie. Na bazie interakcji z rówie
śnikami młode zwierzę musi nauczyć się rozpoznawać osobniki własnego
gatunku, kontrolować agresywność, odróżniać zabawę od zachowań „na serio",
sygnalizowania własnej uległości itd. Socjalizacja u psa domowego była
przedmiotem intensywnych badań (Scott i Fuller, 1965). Wykazano, że klu
czowy jest tu okres między 4. a 12. tygodniem życia, tzw. szeroki okres socjali
zacji. Izolacja w tej fazie może spowodować poważne dysfunkcje behawioru,
jak np. nadpobudliwość na bodźce z otoczenia i zanik umiejętności unikania
bodźców szkodliwych (np. ognia).
Przykładem izolacji w stosunku do określonych bodźców są m i n . ekspery
menty pokazujące wpływ deprywacji na widzenie i związane z nim zachowanie
się. Kiedy kocięta w wieku 2-4 miesięcy są pozbawione normalnych doświad
czeń wizualnych, wytwarza się u nich funkcjonalna ślepota. Upośledzenie to
znajduje odzwierciedlenie w niedorozwoju odpowiednich struktur mózgowych.
Natomiast zmiany takie nie następują, gdy identycznie postępuje się z dorosłymi
osobnikami (Wiesel i Hubel, 1965).
Obraz zmian behawioru, jaki zachodzi w ontogenezie, nie byłby pełny bez
omówienia występującego u niektórych gatunków wpajania, wdrukowywania
albo reakcji piętna (ang. imprinting). Jest to rodzaj szybkiego uczenia się, które
następuje w określonej, zazwyczaj krótkiej fazie po urodzeniu lub wykluciu.

110
Klasyczna forma tego zjawiska, tzw. wpajanie afiliacyjne występuje np. u pta-
ków-zagniazdowników i jest ważnym mechanizmem więzi między potom
stwem a rodzicami. Pisklęta tych gatunków muszą podążać za rodzicami, szybko
zatem powinny nauczyć się przynajmniej jakiejś cechy wyglądu osobników
dorosłych.
Przeprowadzono liczne badania laboratoryjne, które ujawniły interesujące
aspekty tego zjawiska (Hess, 1972). Okazało się, że reakcja piętna jest w dużej
mierze zautomatyzowana. Na przykład, wpajanie afiliacyjne u piskląt kaczki-
-krzyżówki kształtuje się między 13 a 26 godzinami po wykluciu (Grier, 1984).
Jeśli w tej fazie kaczęta zostaną eksponowane na inny obiekt niż przypominający
rodziców (np. na człowieka lub jakiś przedmiot nieożywiony), wtedy to ten
obcy obiekt zostaje wdrukowany i za nim pisklęta podążają. Kolejną interesu
jącą cechą reakcji piętna jest fakt, iż siła wpajania jest tym większa, im większy
wysiłek wkłada pisklę w podążaniu za obiektem, i że sama obecność obiektu,
możliwość widzenia go przez młode ptaki, jest dla nich nagrodą.
Kontrowersyjna jest kwestia nieodwracalności raz ukształtowanej reakcji
piętna. Wpojenie prawidłowe (obrazu osobnika rodzicielskiego) jest nieodwra
calne, natomiast w przypadku obiektów sztucznych, jak się wydaje, istnieje
możliwość przeniesienia reakcji piętna z obiektu bardziej nienaturalnego - np.
z człowieka, na bardziej naturalny - na ptaka innego gatunku (Hess, 1972).
Do zjawisk wpajania afiliacyjnego należy także rozpoznawanie przez rodzi
ców swojego potomstwa po cechach wizualnych (u niektórych ptaków) i po
zapachu (np. u owiec). Jak kształtuje się to zjawisko w przypadku owcy, opisano
wcześniej (patrz rozdz. 3).
Oprócz tworzenia więzi rodziców z potomstwem, podobny do wyżej opisa
nego proces zachodzi wtedy, gdy u zwierzęcia kształtuje się obraz swojego
przyszłego partnera na czas rozrodu. Jest to tak zwane wpajanie seksualne.
Polega ono na przyswojeniu sobie pewnych charakterystycznych cech własnego
gatunku. U ptaków okres, w którym się kształtuje, jest dłuższy niż w przypadku
wdrukowania afiliacyjnego, trwa nawet kilka tygodni - od momentu wyklucia.
I w tym wypadku zwierzę może wpoić sobie nieprawidłowy obiekt (tzw. wdru
kowanie nieprawidłowe - ang. malimprinting). Zjawisko to dość często spotyka
się w ogrodach zoologicznych, gdzie ptaki są odchowywane przez ludzi, nie
mając jednocześnie kontaktu z osobnikami własnego gatunku. W efekcie to
właśnie obraz człowieka wdrukowuje się jako przyszły partner seksualny.
Wpajanie seksualne u ptaków jest bardzo zróżnicowane. Silnie zaznacza
się u niektórych ptaków drapieżnych, u pewnych gatunków żurawi ujawnia się
jedynie pod nieobecność naturalnych partnerów, natomiast u niektórych gatun-

I l l
ków jest praktycznie nieobecne: jeśli nie ma naturalnego partnera, do rozrodu
po prostu nie dochodzi (Grier, 1984).
W literaturze dotyczącej zachowania się zwierząt do wpajania klasyfikuje
się też inne formy szybkiego uczenia się przez młode zwierzęta, których mecha
nizmy są jednak słabiej poznane. Należy tu np. wpajanie topograficzne, utrwala
jące u młodego osobnika obraz ojczystego terenu, wpajanie pokarmowe, czyli
wczesne kształtowanie się preferencji pokarmowych i inne.
Jak widać z powyższego zestawienia, w ontogenezie okresu postnatalnego
dochodzi do wielu zmian zorganizowanych w czasie. Postrzeganie otoczenia,
umiejętności społeczne czy wpajanie zachodzą u zwierzęcia w pewnym, okre
ślonym przedziale wieku, który można eksperymentalnie zidentyfikować. Te
typowe dla pozyskiwania określonych umiejętności okresy rozwoju noszą
nazwę okresów krytycznych (ang. critical period lub sensitive period). Wiedza
o okresach krytycznych u poszczególnych gatunków jest niezbędna nie tylko
dla przyrodników, ale również dla hodowców-praktyków, zajmujących się
odchowem młodych zwierząt.
6.5. Przykładowe zachowanie się - śpiew
ptaków
Wiele z tego, co powiedziano wyżej o determinacji behawioru przez zało
żenia genetyczne i czynniki środowiskowe sumuje się w złożonym zachowaniu,
jakim jest śpiew ptaków. Śpiewają głównie ptaki z rzędu wróblowatych i to
samce. Jak już wspomniano wcześniej, pieśń w odróżnieniu od okrzyku jest
dłuższa i ma rozbudowaną melodią. Jej funkcja związana jest z rozrodem (gło
sowe znakowanie terytorium rozrodczego i faza zalotów). Komplikacja tego
zachowania u ptaków wynika z faktu, że u wielu gatunków występują lokalne
warianty pieśni (tzw. dialekty) oraz ze zdolnością naśladowania odgłosów
osobników jednego gatunku przez przedstawicieli innego.
U niektórych ptaków pieśń ma charakter wrodzony (np. u pasożytów lęgo
wych, których samice składają jaja w gniazdach innych gatunków), u innych
wymaga impulsów ze strony środowiska. W tej ostatniej grupie istnieją gatunki,
u których raz wytworzona pieśń utrzymuje się przez całe życie, i takie, które
„produkują" ją od nowa w każdym sezonie lęgowym. W badaniach nad rozwo
jem śpiewania ptaków posługiwano się różnorodnymi metodami, między in
nymi odchowywano pisklęta-samczyki w izolacji i w pomieszczeniu dźwię-

112
koszczelnym, po czym odtwarzano im z taśmy magnetofonowej nagrane pieśni
osobników tego samego i innych gatunków (Marler, 1970).
Badania te wykazały, że podstawowym warunkiem dla rozwinięcia się pie
śni typowej dla dorosłego osobnika jest usłyszenie jej przez młodego ptaka
w okresie krytycznym. Nie ma on wtedy jeszcze zdolności wokalizowania, ale
zapamiętuje to, co usłyszał, ignorując pieśni obcych gatunków. Drugą, niezwy
kle ważną faząjest jakby ćwiczenie się w śpiewie. Młody ptak próbuje śpiewać,
słyszy własne popisy wokalne, konfrontując je z zapamiętanymi. W efekcie daje
to w końcu gotową pieśń. Jeśli osobnik jest głuchy, wtedy ukształtowanie nor
malnej pieśni jest niemożliwe.
Na tym jednak proces formowania pieśni się nie kończy. W naturze każdy
ptak żyje w określonym środowisku akustycznym i słyszy odgłosy wydawane
przez inne, żyjące obok osobniki własnego gatunku. Ma więc pewne wzorce do
naśladowania. Okazuje się, że u niektórych gatunków, np. szpaków {Sturnus
spp.), występuje elastyczność pozwalająca młodemu samcowi opanować nie
tylko obowiązujący w danym miejscu dialekt, ale kopiować pieśń starszych
i silniejszych osobników, żyjących na sąsiednich terytoriach. Zachowując się
w ten sposób, młody samiec tym samym korzysta prawdopodobnie z rodzaju
ochrony przed ewentualnymi zakusami na terytorium ze strony swoich rówie
śników (Alcock, 1993).

7.
Nauka
o behawiorze
- etologia
7.1. Początki
W 1951 roku ukazała się książka holenderskiego zoologa Nico Tinbergena
„Badania nad instynktem" (The Study of Instinct). Okazała się ona jedną z naj
bardziej wpływowych prac dotyczących zachowania się zwierząt i określała
ramy teoretyczne dla nowej dyscypliny - etologii.
Słowo „etologia" pochodzi od greckiego „ethos" (= obyczaj) i „logos"
(= słowo, rozum). Według najprostszej definicji jest to więc nauka „o obycza
jach, zawierająca badania i opis zachowań zwierząt" (Husson, 1970). Już w tym
bardzo lapidarnym sformułowaniu zawarta jest pewna ważna cecha etologii,
a mianowicie: jej orientacja w kierunku badania spontanicznie manifestujących
się zachowań.
Zaleca się więc obserwacje behawioru, dzięki któremu zwierzę
zaspokaja swoje naturalne potrzeby (np. zdobywanie pokarmu, rozród). Jest to
zachowanie typowe dla gatunku. Zrozumiałe, że taki behawior należy obser
wować przede wszystkim w środowisku naturalnym lub w zbliżonych warun
kach wiwaryjnych (akwarium, terrarium itd.).
Oczywiście tak sformułowany program badań behawioru nie wziął się
z próżni. Patrząc z dzisiejszej perspektywy, tendencje do studiowania zacho
wania się zwierząt w naturze można zaobserwować już od XVIII wieku. Na
przykład, informacje takie gromadził leśnik królewski we Francji i filozof,
Charles Leroy (1723-1789), który wprowadził postulat porównawczego bada
nia zachowania się różnych gatunków.
Okres ekspansji kolonialnej w XIX wieku zaowocował także wyprawami
naukowymi oraz pierwszymi badaniami i eksperymentami terenowymi w zakre
sie zoologii i behawioru, za których pioniera można uznać angielskiego, eks
centrycznego podróżnika Charlesa Watertona (1782-1865). Znaczny wkład do
powstania etologii wniósł Charles Darwin (1809-1882) nie tylko swoją teorią
doboru naturalnego, lecz również porównawczymi studiami nad behawiorem

114
i wyrażaniem emocji (O wyrazie uczuć u ludzi i zwierząt, 1872). Inny Anglosas,
znany zresztą Darwinowi, i wydawca części jego spuścizny - George Romanes
(1848-1894) jako pierwszy podjął próbę naukowego badania inteligencji zwie
rząt, jednak bez utrzymania należytych rygorów metodycznych (Animal Intelli
gence,
1882).
Przeciwstawne, fizjologiczne podejście reprezentował przybyły z Niemiec
do Ameryki Jacques Loeb (1859-1924). Zapoczątkował on badania nad zauto
matyzowanymi, orientacyjnymi reakcjami zwierząt. Obserwując ptaki, na ich
wrodzone, niezmienne gatunkowo zachowania zwrócił uwagę brytyjski badacz
Alexander Spalding. Jako pierwszy odnotował on zjawisko wpajania, które
później przeanalizowali inni badacze. Zoolog o wszechstronnych zainteresowa
niach, Charles Whitman (1842-1910), należy niewątpliwie do amerykańskich
„ojców-założycieli" etologii, który sformułował zasadę, iż cechy zachowania
się podlegają ewolucji tak samo, jak morfologiczne. Jego uczeń, Wallace Craig
(1876-1954), zidentyfikował natomiast ważne dla etologii komponenty zacho
wania - fazę apetencyjną i konsumacyjną.
Europa nie pozostawała w tyle. Szczególne zasługi dla powstania etologii
ma szkoła niemiecka, z Oskarem Heinrothem (1871-1945), dyrektorem akwa
rium w ZOO w Berlinie, lecz z zamiłowania także ornitologiem, który badał
porównawczo zachowanie się ptaków (Ptaki Europy Środkowej, 1926-1933).
Badacz ten nie tylko sporządził inwentarze gatunkowych zachowań, ale zwrócił
także uwagę na te wzorce behawioru, które stały się sygnałami. Francuzi,
w osobie przede wszystkim Jeana Fabre (1823-1915), wsławili się natomiast
badaniami nad biologią owadów, w tym nad zachowaniem się tych zwierząt.
Teoretycznych podstaw dla centralnej w etologii teorii instynktu dały
przede wszystkim prace amerykańskiego psychologa społecznego Williama
McDougalla (1871-1938). To jemu głównie należy zawdzięczać wprowadze
nie w epoce nowożytnej do obiegu myśli psychologicznej pojęcia instynktu,
motywacji, zachowań nastawionych na cel, które były elementami jego teorii
„hormizmu". Wpływ na etologie (a zwłaszcza na rolę popędów) wywarła
również biologizująca teoria psychoanalizy Sigmunda Freuda (1856-1939).
Także idee Niemca, Jacoba Uexkulla (1864-1944), związane z oryginalnym
widzeniem relacji organizm-środowisko, wpłynęły zapewne na teorię bodź
ców kluczowych.
Jak widać z powyższego krótkiego przeglądu, w pierwszym okresie udział
badaczy amerykańskich w formowaniu zrębów etologii był znaczący. Sytuacja
zmieniła się po II wojnie światowej, kiedy to badania w dużej mierze skoncen
trowały się w Europie, a etologia stała się „szkołą europejską". Trzy nazwiska

115
są wymieniane jako najważniejsze ze względu na dorobek badawczy i prace
teoretyczne noszących je osób: Austriacy - Konrad Lorenz (1903-1989) oraz
Karl von Frisch (1886-1982), a także Holender Nicolaus Tinbergen (1907-
-1988). Cała trójka została uhonorowana w 1973 roku Nagrodą Nobla w dzie
dzinie fizjologii, co było także symboliczną nobilitacją etologii i dostrzeżeniem
jej osiągnięć.
Najstarszy spośród trójki, Karl von Frisch, interesował się zmysłowym
postrzeganiem u zwierząt, a w szczególności rolą wzroku i węchu. Ważne miej
sce w jego badaniach zajmowały także eksperymenty dotyczące słuchu u ryb.
Jednak Frisch zdobył międzynarodową sławę dzięki swoim badaniom dotyczą
cym pszczoły miodnej, u której badał postrzeganie barw i węch. Rozszyfrował
on także rolę tzw. tańca pszczół jako swoistej metody komunikowania się mię
dzy tymi owadami (zob. wcześniej, rozdział 3, część dotycząca komunikacji).
Uczeń Oskara Heinrotha, Konrad Lorenz, z upodobaniem prowadził obser
wacje terenowe, badał również rozmaite gatunki zwierząt (przede wszystkim
ptaków), z którymi miał bezpośredni kontakt. Umiejętność obchodzenia się ze
zwierzętami Lorenz mógł zdyskontować, badawczo odchowując od pisklęcia
grupę dzikich gęsi. Dzięki temu mógł on praktycznie nieprzerwanie prowadzić
obserwację behawioru tych w środowisku naturalnym dość płochliwych ptaków.
To właśnie obserwacje spontanicznego zachowania się ptaków wodnych (gęsi
i kaczek), a także przedstawicieli innych gatunków dostarczyły austriackiemu
badaczowi materiału do systematyzacji i wyjaśniania zachowań, a w konse
kwencji do stworzenia wielu koncepcji teoretycznych. Jako teoretyk Konrad
Lorenz ma równie znaczące osiągnięcia co jako badacz terenowy. Jest twórcą
między innymi pomysłowego modelu instynktu. Znana jest jego kontrowersyjna
koncepcja roli agresji w świecie zwierząt i ludzi, pokazał się także jako filozof,
tworząc oryginalną teorię poznania. Konrad Lorenz, człowiek niezwykle
wykształcony, przyrodnik o zacięciu humanistycznym, obdarzony był także
talentem literackim. Świadczą o tym jego książki przeznaczone dla szerszego
grona czytelników, wydane także w języku polskim (np. Tak zwane zło, I tak
człowiek trafił na psa, Opowiadania o zwierzętach).
Także Nicolaus Tinbergen okazał się bardzo płodny zarówno jako badacz
terenowy, jak i teoretyk. Jeszcze przed II wojną światową prowadził obserwa
cje dotyczące orientacji u jednego z gatunków os. Jego ulubionym przedmio
tem obserwacji stały się jednak mewy, którym zresztą poświęcił książkę (The
Herring GulVs World,
1953). Doświadczenia Tinbergena związane z wyzwala
niem zachowań żebrzących u mewy srebrzystej (Larus argentatus), jak też
zalotów godowych u ciernika (Gasterosteus aculeatus) należą do klasycznych

116
prac cytowanych w licznych podręcznikach i opracowaniach. W dużej mierze
na ich kanwie powstały niezwykle ważne dla etologii pojęcia bodźców kluczo
wych i superwyzwalaczy. Wspomniana wyżej praca Tinbergena - Badania nad
instynktem,
może uchodzić za prawdziwą „biblię" etologii wczesnej fazy roz
woju tej nauki. W niej został przedstawiony niezwykle znaczący dla rozwoju
etologii tzw. hierarchiczny model instynktu.
7.2. Klasyczny okres etologii - podstawowe
koncepcje
O orientacji badawczej etologii wspomniano już wyżej. Biorąc pod uwagę
postulat obserwacji zachowań spontanicznych w naturze, nie dziwi też, że mode
lowymi zwierzętami obserwowanymi przez etologów w pierwszej fazie były
przede wszystkim ptaki, ryby i owady. U tych właśnie grup zwierząt wzorce
behawioru typowe dla gatunku ujawniają się w sposób spektakularny i nietrudny
do obserwowania (np. zachowanie godowe). Poza tym zwierzęta te łatwo jest
trzymać w laboratorium i stworzyć im warunki zbliżone do naturalnych.
Jako praktykę badawczą wczesna etologia preferowała obserwację i bardzo
szczególne eksperymenty, w których posługiwano się makietami, mającymi
wyzwalać naturalne zachowania się zwierząt. Zachowania instynktowe anali
zowano szczegółowo, utrwalając je na taśmie filmowej.
Aby poznać cele badań stawiane sobie przez etologów we wczesnym okre
sie, dobrze będzie rozpocząć od przykładu, którym jest zachowanie godowe
ciernika, opisane przez Tinbergena (1965). Ma ono następujące sekwencje
(rys. 10):
1. Wiosną samiec osiąga kondycję ropłodową, która manifestuje się określo
nym ubarwieniem (czerwony brzuch i przezroczystoniebieski grzbiet).
Buduje gniazdo z wodorostów sklejonych specjalną wydzieliną.
2. Samica z jamą brzuszną nabrzmiałą od ikry wpływa na terytorium samca.
3. Samiec wykonuje zygzakowaty taniec.
4. Samica daje się prowadzić do gniazda i wchodzi do niego.
5. Samiec szturcha samicę pyskiem w podstawę ogona.
6. Samica składa ikrę w gnieździe.
7. Samica opuszcza gniazdo, wchodzi do niego samiec i zapładnia komórki
jajowe.
8. Samiec przepędza samicę.
9. Samiec dogląda zapłodnionych jaj (stróżowanie, wentylowanie itd.).

117
R y s u n e k 10. Zachowanie rozrodcze u ciernika (za Tinbergen, 1965, zmodyfikowane): 1 - samiec
pokazuje czerwony brzuch, 2 - widok samicy o ciele nabrzmiałym od ikry, 3 - zygzakowaty taniec
samca i ocieranie się o samicę, 4 - samica przyjmuje pozycję z uniesioną głową, 5 i 6 - samiec
prowadzi samicę do gniazda, 7 - samiec przechyla się na bok, ukazując wejście do gniazda, 8 - samica
wchodzi do gniazda, 9 - samiec uderza pyskiem w ogon samicy, 10 - samica składa ikrę, 11 - samiec
wchodzi do gniazda i zapładnia ikrę

118
W jaki sposób zachowanie to może budzić zainteresowanie badacza o orienta
cji etologicznej? Można to przedstawić w postaci kilku pytań:
1. W jaki sposób w trakcie ewolucji doszło do ukształtowania się takiego
zachowania godowego? Czy u pokrewnych gatunków wygląda tak samo?
Jest to pytanie o ewolucją.
2.
W jaki sposób taki behawior ukształtował się w ciągu życia ciernika? Czy
zygzakowaty taniec wymaga uczenia się? Czy samcowi udaje się doprowa
dzić samicę do gniazda już za pierwszym razem? Jest to pytanie o rozwój
osobniczy.
3. Co jest przyczyną godowego zachowania ciernika? Czy i jakie procesy
zachodzące w organizmie ryby powodują taki behawior? Czy istotne są
zmiany jakiegoś czynnika środowiskowego, np. długości dnia świetlnego?
Jest to pytanie o przyczyną.
4. Jaką rolę dla przeżycia i sukcesu reprodukcyjnego osobników obu płci
odgrywają poszczególne sekwencje zachowań? Na przykład, dlaczego sa
miec szturcha samicę? Dlaczego wentyluje zapłodnioną ikrę? Jest to pytanie
o funkcją.
Spośród wyżej wymienionych zagadnień etologom najistotniejsze wydawało
się pytanie o przyczynę. Interpretowanie zachowania się, wyjaśnianie, jak wła
ściwie zwierzę reaguje na otaczające go środowisko i w jaki sposób dochodzą
do skutku złożone wzorce behawioru, wymagało zidentyfikowania przyczyn.
Powiedzmy na marginesie, że z kolei pytanie o funkcję stało się podstawą
dla omawianej poniżej innej szkoły, ekologii behawioralnej.
Mówiąc najkrócej, podstawowa dla etologii wczesnej fazy teza głosi, że:
Czynnikiem sprawczym zachowań są popędy, które prowadzą do zacho
wań instynktowych. Bezpośrednie wyzwalanie jakiegoś zachowania in
stynktowego zachodzi pod wpływem pochodzących ze środowiska zwierzę
cia bodźców kluczowych, które wywołują określone fazy zachowania się,
czyli koordynacje wrodzone. Proces ten zachodzi przy udziale mechanizmu
wyzwalającego.
Poniżej zamieszczona jest krótka charakterystyka poszczególnych członów
tego twierdzenia.
Popęd
Do zachowania się w określony sposób skłania zwierzę popęd (ang. drive),
zwany inaczej motywacją. Jego działanie można przedstawić jako dążność do
przywrócenia stanu równowagi w organizmie (zaspokojenie potrzeby) lub
uwolnienie się spod działania szkodliwego czynnika. W przypadku popędu
głodu np. nierównowaga jest związana z obniżeniem się poziomu cukru we
krwi. Prowadzi to do dalszych zmian, które w konsekwencji wywołują zacho
wanie się poszukiwania pokarmu. Natomiast klasycznym przykładem uwolnię-
119
nia się spod działania szkodliwego czynnika jest wpływ bólu i towarzysząca
mu reakcja (odsunięcie się od jego źródła). Można powiedzieć, że niektóre
popędy mogą przyciągać zwierzę do atrakcyjnego bodźca, inne natomiast pro
wadzą do usuwania się od bodźca szkodliwego.
Używając przyjętej ogólnie nomenklatury, istnieją więc popędy zachowaw
cze (apetytywne) i ochronne (awersyjne). Osobną, trzecią kategorię stanowi
popęd eksploracyjny, dzięki któremu zwierzę zbiera informacje o otaczającym
środowisku (miejsca rozmieszczenia pokarmu, droga ucieczki z określonego
miejsca itd.). Zaspokojenie popędu eksploracyjnego musi się wiązać z różnymi
doświadczeniami - przyjemnymi i nie, stąd też nie można go zakwalifikować
ani do pierwszej, ani do drugiej spośród wymienionych niżej kategorii.
Nie wszystkie popędy ujawniają się jako gotowe (odziedziczone). Niektóre
z nich kształtują się w wyniku doświadczeń osobnika. Płynie stąd kolejny podział
na popędy pierwotne i wtórne. Na przykład, reakcja na ból jest wynikiem
popędu pierwotnego, natomiast strach przed bólem to popęd wtórny.
Klasyfikacja popędów oparta na wymienionych powyżej podziałach zawarta
jest w tabeli 14.
Tabela 14. Klasyfikacja popędów (za Beck, niepublikowane, za pozwoleniem autora)
Wyszczególnienie
Popędy pierwotne
Popędy zachowawcze
(apetytywne)
Popędy ochronne
(awersyjne)
Popęd eksploracyjny
Wyszczególnienie
Popędy pierwotne głód
pragnienie
popęd seksualny
popęd macierzyński
popęd agresji
ból
strach wrodzony
popęd ucieczki
-
Popędy wtórne
więź w parze
więź matka - potomstwo
więź seksualna
strach przed bólem
frustracja
-
Bardzo często zdarza się, że w określonym momencie w organizmie wzbu
dzony jest jednocześnie więcej niż jeden popęd (np. głód i pragnienie). Możliwe
są wówczas trzy interakcje popędów: torowanie, hamowanie i antagonizm.
W pierwszym przypadku jeden popęd ułatwia wzbudzanie innego, np. głód toru
je agresję. W drugim - następuje redukowanie jednego z popędów przez drugi,
np. seksualnego przez głód. Z punktu widzenia zachowania się najciekawszy jest
trzeci przypadek, kiedy na zwierzę działa jednocześnie popęd apetytywny i awer-
syjny. Silniejszy popęd może przeważyć. Ale gdy równoważą się one ze sobą,
wtedy wywoływane przez nie zachowania wzajemnie się niwelują. W wyniku
takiej interakcji dochodzi często do pojawienia się specyficznych zachowań ago-
nistycznych, jak przerzutowe czy przenoszone (zob. rozdz. 3).

120
Przedstawiony wyżej schemat jest uproszczony i nie uwzględnia pewnych interesujących warian
tów zachowania, które można przewidzieć przy pojawianiu się dwóch stanów popędowych. Co
stanie się np., gdy pies jest jednocześnie i głodny, i spragniony, ale popęd związany z głodem jest
jedynie w niewielkim stopniu silniejszy. Jeśli założymy, że pokarm i woda są łatwo dostępne, ale
usytuowane w różnych miejscach w pomieszczeniu laboratoryjnym, można teoretycznie przewi
dzieć następujący scenariusz wydarzeń:
Zgodnie z założeniem redukcji jednego popędu przez drugi, pies zbliży się do miski z pokar
mem i zacznie jeść. W trakcie jedzenia jednak j e g o motywacja pokarmowa obniży się i górę weźmie
chęć do zaspokojenia pragnienia. Pies porzuci więc pokarm i przejdzie do miski z wodą. Z kolei
w trakcie zaspokajania pragnienia poczuje głód, gdyż teraz nieco silniejsza jest motywacja pokar
mowa. Teoretycznie więc w opisanej wyżej sytuacji pies powinien nieustannie krążyć między
miskami zawierającymi pokarm i wodę.
O c z y w i ś c i e w praktyce do tego nie dochodzi. Gdyby scenariusz rozwijał się w sposób opisany
powyżej, zwierzę więcej czasu spędzałoby na krążeniu między miskami niż najedzeniu i piciu. Tak
w i ę c zagadnienie zaspokajania popędu splata się tu z problemem wyboru i trwałości zachowania się
zwierząt. Badacze przypuszczają, iż w analizowanej sytuacji można podać dwa wyjaśnienia trwało
ści zachowań pokarmowych. Przede wszystkim, im bardziej pies zbliża się do miski z pokarmem,
1 tym bardziej zwiększa się siła bodźca pokarmowego, która „trzyma" zwierzę i pozwala poświęcić
mu więcej czasu na jedzenie. Innym wyjaśnieniem jest przyjęcie istnienia hipotetycznego „systemu
sytości", który włącza się, hamując - powiedzmy - pobieranie pokarmu w momencie, gdy pies jest
nasycony, a nie jedynie wówczas, gdy popęd głodu uległ choćby nawet znacznej redukcji (Balke-
nius, 1995). _ _ _ _
=
_ ^
=
^ ^ ^
=
^ _
Instynkt i koordynacja wrodzona
Zaspokojenie popędu następuje w wyniku zachowania instynktowego.
Instynkt był centralnym pojęciem etologii w fazie „klasycznej", aczkolwiek
także pojęciem kontrowersyjnym, różnie rozumianym przez rozmaitych badaczy
(zresztą również przez nieetologów). Immelhmann i Beer (1989) przytaczają
następujące wersje rozumienia tego terminu w literaturze psychologicznej:
• działanie celowe bez przewidywania skutków i bez wcześniejszego
uczenia się;
• siła napędowa do spełniania jakichś biologicznych funkcji, jak migra
cja czy rozmnażanie się;
• niezmienny i typowy dla gatunku wzorzec działania;
• genetycznie uwarunkowany i sterowany wewnętrznie mechanizm
wpływający na zachowanie charakterystyczne dla gatunku.
Natomiast w ujęciu etologów instynkt był pojmowany jako ukształtowany
ewolucyjnie, wrodzony i zasadniczo niezmienny mechanizm zachowania się,
oparty na popędzie. Jednak przyznać trzeba, że w tej kwestii istniały pewne
różnice ujęcia nawet między K. Lorenzem i N. Tinbergenem, co uwidaczniało
się w różnych prezentowanych przez nich modelach instynktu (model psycho-
hydrauliczny Lorenza i hierarchiczny Tinbergena).

121
Podstawową jednostką zachowania instynktowego jest koordynacja wrodzo
na (ang. fixed action pattern, skr. FAP), mniej lub bardziej złożony wzorzec
ruchowy, jednak o wyraźnie większej komplikacji od prostego odruchu. Koordy
nacja wrodzona z dużą dokładnością powtarza się u każdego normalnego osobni
ka danego gatunku i nie wymaga doświadczenia (od razu manifestuje się w peł
ni). Kolejnymi cechami koordynacji wrodzonej jest to, iż jest wyzwalana przez
specyficzne bodźce i ma przynależną sobie energię (o czym niżej). Jest to także
rodzaj zachowania „spustowego" (ang. triggered), który raz wyzwolony trwa
nadal, nawet gdy zmieniające się warunki otoczenia czynią go bezużytecznym.
Wywołane przez popęd zachowanie może składać się z kilku koordynacji
wrodzonych, następujących po sobie w określonej kolejności. Omówione wcze
śniej zachowanie godowe samca ciernika jest właśnie takim ciągiem stopniowo
manifestowanych koordynacji wrodzonych.
Bodziec kluczowy
Koordynacja wrodzona zostaje wyzwolona przez specyficzne bodźce po
chodzące z otoczenia, tzw. bodźce kluczowe (ang. sign stimuli, key stimuli).
Bodźce te należą w zasadzie do wszystkich modalności sensorycznych,
występują więc jako: wizualne, akustyczne, zapachowe i dotykowe.
W omawianym wcześniej zachowaniu się ciernika bodźcami kluczowymi
wizualnymi inicjującymi behawior godowy są np. czerwona barwa brzucha
samca i widok napęczniałego od ikry brzucha samicy. Typ bodźca akustycznego
to np. specyficzne kwilenie indycząt, które wyzwala zachowanie opiekuńcze
indyczki (głucha indyczka może zadziobać swoje pisklęta!). Natomiast przy
kładem bodźców węchowych może być, omawiane już wielokrotnie w innych
miejscach, działanie feromonów u owadów. Bodziec kluczowy występuje jako
pojedynczy lub stanowi konfigurację sumujących się bodźców. Na przykład,
w zachowaniu terytorialnym samiec ciernika wykazuje większą agresywność
w stosunku do obcego osobnika, który przyjmuje spionowaną pozycję grożącą
- głową w dół. Sumuje się tu efekt działania dwóch bodźców wizualnych -
barwy czerwonej i pozycji ciała (Manning, 1972).
Z tego, co powiedziano wyżej, wynika, iż bodźce kluczowe mogą wystę
pować w kontekście społecznym (zachowania godowe, terytorialne itd.) i poza
nim (np. bodźce w relacji drapieżnik - ofiara). Te, które występują w kontekście
społecznym, określane bywają niekiedy jako wyzwalacze (ang. releasers).
W behawiorze godowym u ciernika zachowania obu osobników pary stają się
wizualnymi wyzwalaczami dla behawioru drugiego osobnika. I tak np. zygza
kowaty taniec samca ciernika wyzwala u samic podążanie do gniazda, a zajęcie
przez samicę miejsca w gnieździe powoduje wyzwolenie reakcji uderzanie
pyskiem przez samca.

122
Z zagadnieniem wyzwalania koordynacji wrodzonych związana jest także
koncepcja ponadnormalnych bodźców kluczowych albo superwyzwalaczy
(ang. supernormal stimuli, super releasers). Poprzez sztuczne nasilenie cechy
bodźca kluczowego uzyskuje się efektywniejsze wyzwolenie koordynacji wro
dzonej. Przykładem są doświadczalnie wykazane zachowania niektórych pta
ków, które zaniedbują wysiadywanie własnych jajek dla opieki nad znacznie
większym jaj em-atrapą, podrzuconym przez eksperymentatorów (rys. 1 la).
a b
R y s u n e k 1 1 . Przykłady superwyzwalaczy: a - duża makieta jaja ostrygojada wyzwala u ptaka
zachowanie polegające na próbach wtaczania do gniazda przy jednoczesnym ignorowaniu
autentycznych jaj, b - plamy na skrzydłach motyli (na górze - Taenaris catops, na dole - Caligo
illioneus) jaskrawo odcinające się od tła w przejaskrawionej postaci przypominają ocz y ptaków
drapieżnych - co ma odstraszać małe ptaki (wg różnych źródeł)
Istnieje hipoteza, że superwyzwalacze mogą pełnić także ważną rolę
w życiu zwierząt. Na przykład, barwny, jaskrawo odcinający się, kolisty deseń
na skrzydłach motyla miałby imitować oczy ptaków drapieżnych, a tym samym
działać odstraszająco na prześladowców owadów - małe ptaki (rys. 1 lb). Wia
domo również, iż efekt superwyzwalaczy wykorzystują ptaki stosujące strategię
pasożytowania lęgowego. Na przykład, jaja kukułki (Curculus canorus), które
składane są w gniazdach innych ptaków, są zwykle większe i jaskrawo ubar-

123
wionę. Powoduje to, iż przybrani rodzice zajmują się przede wszystkim inku-
bowaniem ich, a nie własnych.
Prawo heterogenicznego sumowania bodźców
Prawo to jest uogólnieniem obserwacji dotyczących bodźców kluczowych.
Głosi ono, iż wyzwolenie zachowania instynktowego uzależnione jest od sumy
bodźców kluczowych wyzwalających ten behawior, ale których ważność dla
organizmu może ulegać relatywnej zmianie. Tak więc jeżeli np. wyzwalanie
zachowania ptaków względem jaj uzależnione jest od wielkości i plamistości
tych ostatnich, to zanik plam może zostać zrekompensowany większymi wymia
rami jaj.
Wrodzony mechanizm wyzwalający
Bodźce kluczowe (w najszerszym sensie, łącznie z wyzwalaczami) można
porównać do wiązki wrażeń zmysłowych, które zostały wyselekcjonowane
z szerokiego spektrum bodźców docierających do narządów zmysłowych zwie
rzęcia. W jaki sposób doszło do takiej selekcji?
Według etologów w układzie nerwowym funkcjonuje mechanizm, nazwany
przez nich mechanizmem wyzwalającym albo wrodzonym mechanizmem wy
zwalającym (ang. Innate Releasing Mechanism, skr. IRM). Jego działanie pole
ga na filtrowaniu bodźców. W skrócie wygląda to następująco: dochodzący do
narządu zmysłowego bodziec jest porównywany z zakodowanym genetycznie
wzorcem bodźca kluczowego. Kiedy występuje zgodność, znoszona jest blokada,
która przedtem uniemożliwiała ujawnienie się koordynacji wrodzonej. Na
poziomie anatomofizjologicznym filtrujące działanie mechanizmu wyzwalają
cego związane jest najprawdopodobniej zarówno z aktywnością struktur narzą
dów zmysłów, jak i centralnego i obwodowego układu nerwowego.
Energia specyficznego działania
Niektórzy etolodzy utrzymywali, że każda koordynacja wrodzona ma pewien
przynależny sobie zasób energii, który wyładowuje się, kiedy zwierzę wykazu
je zachowanie instynktowe. Po wyczerpaniu się energia ta, nazwana energią
specyficznego działania (ang. action-specific energy), kumuluje się na nowo. Tak
więc łatwość, z jaką zostaje wyzwalana koordynacja wrodzona, zależy w danym
momencie nie tylko od obecności bodźców kluczowych, ale również od zasobu
energii specyficznego działania.
Jeśli na zwierzę przez długi czas nie oddziałują żadne bodźce kluczowe,
energii tej gromadzi się tak wiele, że uwalnia się ona spontanicznie w postaci
tzw. zachowań upustowych (ang. vacuum activity). Przykładem może być zacho-

124
wanie niektórych ptaków, które wykonują ruchy chwytania owadów, który
w istocie nie ma (Manning, 1972).
Z drugiej strony, nawet przy mniejszej energii zachowanie może b;
wyzwolone, jeśli bodźce kluczowe są wystarczająco silne. Jest to wytłumacz
nie dla zachowania drapieżników określanego jako „nadmierne zabijanie" (an
surplus killing).
Kiedy łasica (Mustela nivalis), albo inny drapieżnik, dostaje s
do kurnika, zabija często więcej niż może zjeść. Dzieje się tak, ponieważ dzi
łanie bodźców kluczowych (w popłochu uciekające ptaki) wyzwala silniejs:
zachowanie polegające na chwytaniu i zabijaniu.
Całościowy obraz zachowania instynktowego
Zależności dotyczące wpływu podstawowych czynników na behawii
zostały uogólnione w postaci zasady podwójnej kwantyfikacji. Mówi ona, ;
wyzwolenie zachowania instynktowego uzależnione jest łącznie od siły popęc
(czynnik wewnętrzny) i bodźców kluczowych (czynnik zewnętrzny).
Przebieg zachowania instynktowego wygląda następująco:
Pierwszym jego etapem jest faza apetencyjna (ang. appetetive), w któr
zwierzę poszukuje stymulacji, czyli bodźców kluczowych. Rozpoczyna się 01
wraz z pojawieniem się określonego popędu. Przykładem może być głodne zwi
rzę, które wyrusza w poszukiwaniu pokarmu. W fazie apetencyjnej często możi
wyróżnić kilka etapów, które wyzwalane są przez odpowiednie bodźce kluczow
Na przykład, drapieżnik polujący na gryzonie słyszy najpierw szmer w trawi
który prowadzi go w określone miejsce, później widzi nieznaczne ruchy traw
pozwalające mu na bliższą lokalizację przestrzenną ofiary i podkradanie się it
Zwierzę przechodzi więc z jednego etapu do drugiego, przy czym pojawienie s
kolejnego bodźca kluczowego hamuje działanie poprzedniego. W ten sposc
powstaje cały łańcuch etapów przygotowawczych (Sadowski i Chmurzyńsl
1989). W obrębie fazy apetencyjnej może wystąpić koordynacja wrodzona, a
z reguły jest to bardziej plastyczna część zachowania instynktowego, podatna r
modyfikację wynikającą choćby z uczenia się (zob. niżej).
W końcu zwierzę odnajduje bodziec, który wyzwala finalną fazę behawioi
instynktowego. Nosi ona nazwę konsumacyjnej, czyli spełniającej (ang. consun
matory act).
W skład fazy konsumacyjnej wchodzi zachowanie sztywne i nii
zmienne, czyli typowa koordynacja wrodzona. Drugim elementem tej fazy je
składowa orientująca - powodująca właściwe ustawienie się zwierzęcia względei
obiektu, który jest jego celem. Na przykład, przed zabiciem ofiary (które jest k<
ordynacją wrodzoną) drapieżnik musi ją w odpowiedni sposób chwycić zębami.
W złożonych zachowaniach instynktowych pojęcie apetencji i konsumac
może być w praktyce dość płynne i zależeć od motywacji. Wykazały to badam
Leyhausena (1960) na kotach domowych i dzikich. Badacz ten stwierdził, i

125
takie wzorce, należące do instynktowego zachowania pokarmowego, jak chwy
tanie, zabijanie, skubanie piór i sierści oraz zjadanie, są relatywnie niezależne
od siebie. Tak więc w określonej sytuacji, jeśli zwierzę jest wystarczająco syte,
zachowaniem konsumacyjnym dla kota może być np. tylko faza chwytania,
zabijania albo skubania, nie zaś zjadanie ofiary.
Warto dodać także, iż zachowania instynktowe, należące do tej samej grupy,
mogą wykazywać różną względną długość fazy apetencyjnej i konsumacyjnej.
Na przykład, w zachowaniu pokarmowym ssaki kopytne mają krótką fazę apeten-
cyjną natomiast długą - konsumacyjną. U ssaków drapieżnych jest odwrotnie.
Model instynktu według N. Tinbergena
Według koncepcji Tinbergena (1951) instynkt odpowiada systemowi hie
rarchicznemu znajdujących się w układzie nerwowym wrodzonych mechani
zmów wyzwalających i ośrodków motorycznych, które powodują stopniowe
ujawnienie się poszczególnych zachowań (ich faz apetencyjnych i konsumacyj-
nych). Za przykład może posłużyć instynkt rozrodczy u samca ciernika.
Mechanizm hierarchiczny tego instynktu zostaje uruchomiony dzięki sty
mulacji natury fizjologicznej (hormonalnej). Wtedy następuje poszukiwanie
przez zwierzę stosownego terytorium (faza apetencyjna). Dopiero, gdy ryba
znajdzie stosowny teren (o czym informuje ją odpowiedni bodziec kluczowy),
mogą zostać odblokowane inne ośrodki, sterowane przez wrodzone mechani
zmy wyzwalające i odpowiadające za zaloty, budowę gniazda, walkę o teryto
rium itd. Jeśli ciernik natrafi na odpowiednie bodźce kluczowe, uruchamia się
program zachowań instynktowych związanych z budową gniazda.
Dalej, w zależności od napotkanych bodźców kluczowych, ciernik będzie
albo zalecał się do napotkanej samicy, albo walczył z konkurentem. Po zalotach
odblokowują się dalsze ośrodki dyspozycyjne zachowań dotyczące opieki nad
ikrą (też oczywiście wyzwalane przez odpowiednie bodźce kluczowe). I tak do
osiągnięcia ostatecznego celu, jakim jest odchów potomstwa.
7.3. Późniejsze zmiany
Patrząc na późniejsze zmiany, które nastąpiły w etologii, można dojść do
wniosku, że zarówno w sferze praktyki badawczej, jak i teoretycznej modyfi
kacji uległo niemal wszystko. Przedmiotem badań stały się już nie tylko małe
zwierzęta o wyrazistych „instynktach", ale także większe ssaki (przede
wszystkim naczelne, drapieżniki i kopytne), liczne ptaki i inne kręgowce.

126
Wprowadzono długotrwałe obserwacje terenowe behawioru dużych ssaków,
badania, których pionierem był Amerykanin George Schaller. Przedmiotem
zainteresowania stały się także zwierzęta domowe, mimo iż z punktu widze
nia klasycznej etologii dysponowały one w pewnej mierze „nienaturalnym"
zachowaniem się.
Stałe doskonalenie metodyki etologicznej wynikało przede wszystkim
z konieczności analizowania coraz większej ilości danych, co spowodowało
formalizację, a nawet pewną matematyzację etologii. Tak jak w innych dzie
dzinach wiedzy, przełomem w analizie danych było szerokie wprowadzenie
techniki komputerowej. Dziś badacz-etolog ma do dyspozycji specjalistyczne
oprogramowanie, które pozwala mu na znacznie szybszą analizę trudnych do
opracowania danych, np. materiału nagranego techniką wideo.
W sferze teoretycznej największa zmiana polegała na otwarciu się etologów
na inne dyscypliny wiedzy, zajmujące się behawiorem. Okazało się, że istnieją
liczne punkty wspólne etologii z endokrynologią, neuropsychologią, ekologią,
genetyką, a także socjologią, antropologią i psychiatrią. Wymiana idei okazała
się płodna. Powstały dyscypliny graniczne, jak np. neuroetologia, badająca
nerwowe podłoże manifestowania się i kontroli zachowań. Nastąpiło też zbliże
nie z behawioryzmem (zob. niżej), początkowo opozycyjną względem etologii
szkołą badawczą.
Analizując zmiany etologii w planie bardziej szczegółowym, można dostrzec,
że znacznej modyfikacji uległo rozumienie instynktu i zachowania instynkto
wego. Przede wszystkim zmienił się pogląd na rolę czynników środowisko
wych w tych zachowaniach. Okazało się, że np. wrodzony mechanizm wyzwa
lający (selekcjonujący bodźce z otoczenia) może być podatny na doświadczenie
osobnicze, co udowadniają eksperymenty dotyczące wpajania (zob. rozdz. 6),
lecz nie tylko (zob. tekst w ramce).
W klasycznym doświadczeniu N. Tinbergen udowodnił, jakie wyzwalacze odgrywają rolę
w relacjach między m e w ą srebrzystą a jej potomstwem. Okazało się, że na żebranie piskląt o po
karm, czyli na ich uderzanie dziobami w dziób matki wpływ miały takie cechy, jak: kształt, barwa
dzioba oraz kontrast pomiędzy j e g o barwą a znajdującą się na nim ciemną plamką (Tinbergen,
1951). Doświadczenie to prowadzono przy użyciu makiet odtwarzających realistycznie głowę
dorosłej mewy.
W trakcie dalszego badania pisklętom pokazywano makietę g ł o w y m e w y srebrzystej i m e w y
śmieszki (Lanis ridibundas). Okazało się, że w wieku 1-2 dni pisklęta nie różnicują makiet i dzio
bią w nie bez większej różnicy. Jednak w miarę upływającego czasu selektywność reakcji wobec
modelu własnego gatunku zaczęła zdecydowanie wzrastać (Hailman, 1967). Świadczy to o zmianie
mechanizmu wyzwalającego się pod wpływem indywidualnych doświadczeń ptaków.

127
Wobec tego dziś mówi się o dwóch typach mechanizmów wyzwalających
- zamkniętym i otwartym. Ten drugi to właśnie mechanizm podatny na wpływy
środowiska. Wiadomo, iż zmiany zachodzą także w fazie apetencyjnej, gdzie
zwierzęta uczą się poprzez oba typy warunkowania (zob. rozdz. 9) i naśladow
nictwo.
Wpływ otoczenia na zachowania instynktowe można też było prześledzić
w aspekcie rozwojowym. Przykładem mogą być teoretycznie najsztywniejsze
formy behawioru - odruchy, wymagające często jednorazowej ingerencji czyn
nika środowiskowego. Na przykład, odruch oddawania moczu u noworodków
szczurów (stanowiący część szerszego zachowania instynktowego) musi być
poprzedzony jednorazowym bodźcem dotykowym ze strony matki. Po takim
pobudzeniu później odruch ten przebiega już sprawnie (Manning, 1972).
Poważnym problemem stało się to, że instynkt wciąż uwikłany był w termi
nologicznej niejasności, a krytycy etologii uznawali je za swoiste „słowo-
-wytrych", które stanowi wystarczające wyjaśnienie zachowania się. Termin
„instynkt" nabrał zresztą negatywnego znaczenia poza obszarem nauk przyrod
niczych. Dlatego też pojawiły się głosy, by z niego zrezygnować.
Także i inne elementy klasycznej teorii etologicznej stały się przedmiotem
dyskusji i krytyki. Na przykład, energię specyficznego działania uznano za
kategorię czysto myślową, nie mającą odniesienia do rzeczywistości. Prawo hete
rogenicznego sumowania bodźców okazało się prawdziwe w stopniu ograni
czonym. Wyszło bowiem na jaw, że w wypadkach niektórych zachowań nie
można mówić o prostym sumowaniu wywołujących je bodźców (Immelhmann
i Beer, 1989). Liczne spory wzbudziły też tylko częściowo omówione tu modele
instynktów, a przede wszystkim najważniejsze - Lorenza i Tinbergena. W sumie,
ze względu na koncentrowanie się na opisie i analizowaniu mechanizmów,
etologom zarzucano zbyt płytkie podejście do problemu zachowania i częste
dawanie nie satysfakcjonujących wyjaśniań wobec złożonych problemów, które
napotykali. „Wiele opisów przypominało raczej etykietki zachowań niż ich
objaśnienie" (Buss, 1999). Stałym przedmiotem krytyki był fakt niepoświęcania
należytej uwagi funkcjonalnemu aspektowi behawioru.

8.
Wykorzystanie
etologii
w praktyce
Podstawowym celem etologii jest zrozumienie zachowania się organizmów.
Jednak badanie przyczyn i mechanizmów behawioru u zwierząt, które mają dla
człowieka jakiekolwiek znaczenie (ekonomiczne, psychologiczne, medyczne
itd.) powinno zarazem prowadzić (jak w przypadku innych nauk) do praktycz
nego wykorzystania zgromadzonej wiedzy.
Poniżej przedstawiono kilka zaledwie przykładów aplikacji odkryć etologów
w różnych sferach życia i działalności człowieka. Wykorzystano wiedzę etolo-
giczną dotyczącą głównie roli bodźców kluczowych (wyzwalaczy), a także
zachowań instynktowych (behawioru specyficznego dla gatunku). Warto też
zauważyć, że w niektórych przypadkach do pilnego wykorzystania wiedzy eto-
logicznej człowiek został wręcz zmuszony, korygując własne błędy w chowie
zwierząt i postępowaniu z nimi.
1. Środowisko naturalne
W nowoczesnej gospodarce zasobami naturalnymi istnieje czasem koniecz
ność manipulowania zwierzętami, regulowania wielkości ich populacji, prze
siedlania pewnej liczby osobników itd. Specjalne działania muszą być często
podejmowane na terenach ustawowo chronionych.
W afrykańskich parkach narodowych zwierzęciem przysparzającym wiele
problemów jest słoń. Chodzi nie tylko o ochronienie go przed kłusownikami,
ale także o pojawiającą się czasem konieczność przesiedlania części osobni
ków, gdy jest ich za dużo na terenie danego parku narodowego. Do takiej sytu
acji doszło właśnie w RPA, gdzie osierocone, młode słonie zostały przesiedlone
z Parku Narodowego Krugera do Pilanesburga. Mimo że w nowym miejscu
zaadaptowały się dobrze, okazało się wkrótce, iż stwarzają nowy, poważny
problem. Młode samce słoni atakowały i zabijały nosorożce białe (Ceratothe-
rium simum).
W latach 1981-1997 zginęło z tego powodu ponad 40 tych równie
rzadkich co słonie i tępionych przez kłusowników zwierząt (Slotów, 2000).
Dlaczego tak się stało? Otóż nie przewidziano możliwego scenariusza, który
wynika ze specyfiki zachowania się słoni. Samce tego gatunku jako nieliczne
wśród ssaków przechodzą co roku okres, który jest odpowiednikiem rui u samic

129
innych gatunków. Ten okres pobudzenia nosi nazwę „musht" i charakteryzuje
się podwyższonym poziomem testosteronu we krwi oraz obecnością charakte
rystycznej wydzieliny z gruczołów skroniowyh. Samce słoni, będące w musht,
stają się bardzo agresywne i toczą ze sobą walki.
Nie uświadomiono sobie, że w normalnych warunkach występowanie pobu
dzenia i jego objawy u młodych słoni są tłumione przez obecność starszych
osobników, które „temperują" agresywność młodszych, zwyciężając je w wal
kach. Przegrana walka powoduje bowiem rychłe zahamowanie objawów
musht. Natomiast pod nieobecność starszych samców pobudzenie: po pierwsze
- pojawiło się wcześniej niż normalnie (tj. w wieku 25-30 lat), po drugie -
trwało ono długo (nawet ponad 130 dni), po trzecie - posiadające potężną ener
gię młode samce wyładowywały agresywność w atakach na osobniki innego
gatunku.
Toteż nic dziwnego, że sytuacja w Pilensburgu poprawiła się bardzo szybko
dopiero wtedy, gdy w 1998 roku skorygowano błąd i sprowadzono tam sześć
starszych samców. Ataki na nosorożce wkrótce ustały.
2. Uprawy i las
Walka człowieka ze szkodnikami upraw trwa właściwie od początków
ludzkiej aktywności rolniczej. Wiek XX charakteryzował się w tym względzie
szerokim zastosowaniem chemii. Jednak środki chemiczne do tępienia owadów-
-szkodników upraw i lasów są z reguły nieprzyjazne dla środowiska naturalnego,
a poza tym po pewnym czasie mogą stracić swoją skuteczność (następuje
uodpornienie się na nie szkodników). Toteż zdecydowanie bardziej pożądana
jest w praktyce rolniczej i leśniczej tzw. kontrola biologiczna, w której również
znajduje zastosowanie wiedza o zachowaniu się zwierząt.
Ogromne znaczenie w takiej formie zwalczania owadzich szkodników mają
feromony. Niektóre spośród tych specyficznych dla danego gatunku i wydzie
lanych naturalnie substancji chemicznych zostały poznane, wyizolowane i użyte
jako swoiste środki „owadobójcze". Feromony u owadów, jak wiadomo, służą
przede wszystkim do wabienia osobnika drugiej płci w czasie rozrodu. Tak
więc można je wykorzystać jako przynętę w rozmaitych pułapkach. Eliminuje
się w ten sposób osobniki jednej płci, co w dalszej perspektywie prowadzi do
spadku liczebności szkodnika.
W jednej z najbardziej masowych tego typu akcji, przeprowadzonej pod
koniec lat 70. XX wieku w Norwegii, posłużono się zsyntetyzowanym feromo
nem do zwalczania korników (Ips spp.). Problem z tymi chrząszczami polega na
tym, iż wydzielane przez nie substancje wabiące informują o pokarmie i przycią
gają zarówno samce, jak i samice. W Norwegii, w omawianym okresie, z racji
inwazji korników wystąpiły ogromne straty w drzewostanie jodłowym, toteż

130
należało podjąć zdecydowane działanie. W akcji zwalczania niszczących drzewa
szkodników posłużono się milionem pułapek przywabiających, w które w ciągu
kolejnych lat złapało się od 2,9 do 4,5 mld korników. W dużej mierze zażegna
ło to niebezpieczeństwo (Marchlewska-Koj, 1988).
Jednak szkodniki upraw to nie tylko owady, lecz także kręgowce. Na przy
kład, dziki królik (Oryctolagus cuniculis) czy zając (Lepus europaeus) potrafią
być destrukcyjne w ogrodach i sadach, niszcząc roślinność uprawną oraz drzewa.
W odstraszaniu tych nieproszonych gości wykorzystuje się ich obawę przed
naturalnymi drapieżnikami, która prawdopodobnie w przypadku wielu mniej
szych ssaków ma charakter wrodzony (Marchlewska-Koj, 1988). Zapach lisa
czy kota (wprowadzony w postaci próbki lisiego tłuszczu, kociego żwirku itd.)
szybko powoduje zamierzony efekt. Podobnie na niektóre gryzonie i króliki
oddziałują odstraszająco bodźce niepokojące i nieznane, np. wkopana w ziemię
pusta butelka, wydająca dźwięki przy podmuchach wiatru. Zbliżoną strategię
stosuje się wobec ptaków, gdzie najskuteczniejszymi „strachami" okazują się
być powiewające na wietrze, falujące tkaniny, zawieszone paski lśniącej folii,
pocięte torby plastikowe itd.
3. Miasto
Wiele gatunków zwierząt dzikich - ssaków i ptaków żyje w dużych mia
stach, korzystając z obfitości pokarmu, który mimowolnie dostarcza im czło
wiek (np. na wysypiskach śmieci). Niektóre zwierzęta znajdują także wspaniałe
kryjówki, zwłaszcza na terenach podmiejskich, w dużych ogrodach itd. Fauna
miejska Ameryki Północnej i Europy obejmuje np. gatunki takie, jak: lis pospoli
ty, kojot, szop pracz (Procyon lotor), niektóre ptaki śpiewające i drapieżne oraz
gryzonie. Osobną grupę stanowią zwierzęta, które „powróciły do natury" -
zdziczały kot, pies i gołąb.
Wpływ wielu spośród tych gatunków nie jest obojętny dla człowieka, są
one bowiem nosicielami groźnych drobnoustrojów chorobotwórczych i pasoży
tów. Problem stanowi m.in. koncentracja odchodów ptaków. W niektórych
miastach mewy (Larus spp.) zadomowiły się na tyle, że ich obecność zagroziła
zanieczyszczeniem komunalnych ujęć wody. Taka sytuacja miała np. miejsce
w Glasgow, gdzie gdy zawiodły inne metody, do odpędzania mew wykorzystano
ich wrodzoną reakcję na sygnał zwiastujący niebezpieczeństwo. Z głośników
umieszczonych w „strategicznych" miejscach emitowano nagranie okrzyku
mewy schwytanej przez drapieżnika. Odgłos ten nadawano, gdy ptaki próbowa
ły lądować w okolicy zbiornika i czyniono to nieregularnie (chodziło o to, by
mewy nie przyzwyczaiły się do sygnału i nie zaczęły go ignorować). Zastoso
wane środki dały rezultat, ponieważ mewy definitywnie przeniosły się w inne
miejsce (Huntingford, 1984).

131
4. Ogród zoologiczny
Współczesny ogród zoologiczny coraz częściej przestaje być zwykłym
zwierzyńcem, prezentującym pewną kolekcję faunistyczną, staje się natomiast
ośrodkiem, w którym (dzięki hodowli) próbuje się chronić gatunki zagrożone
wyginięciem.
Na skutek nacisku opinii publicznej i lepszego rozumienia potrzeb zwierząt
dzikich trzymanych w ZOO próbuje się dla nich tworzyć jak najbardziej kom
fortowe warunki ekspozycji. Nowa nauka - „zoobiologia" zajmuje się właśnie
problemami adaptacji do warunków wiwaryjnych i próbuje określić optimum
środowiskowe dla każdego gatunku. W wielu współczesnych ogrodach już
istniejące ekspozycje, nie odpowiadające potrzebom organizmu zwierzęcego,
często próbuje się udoskonalić poprzez wprowadzanie tak zwanych wzbogaceń
(ang. enrichment). Są to wszelkiego typu elementy, które zwalczają nudę zwie
rząt i działają na nie aktywizująco (np. urozmaicenia wystroju wybiegu, przed
mioty do manipulowania czy urządzenia do wyzwalania aktywności łowieckiej).
Nie ulega wobec tego wątpliwości, że ważnym elementem „zoobiologii" są
badania zachowania się zwierząt, i to zachowań instynktowych.
O tym, że wiedza dotycząca behawioru zwierząt może być niedostatecznie
wykorzystana przekonuje przykład gerenuka (Litocranius walleri). Jest to gatu
nek afrykańskiej gazeli, żyjącej w suchym środowisku (półpustynie). W jednym
z ogrodów zoologicznych wystąpiły kłopoty z rozmnażaniem gerenuków, mimo
iż dysponowano osobnikami w odpowiednim wieku i kondycji. Badania ujaw
niły istotę problemu.
Gerenuk jako zwierzę przystosowane do chronicznego braku wody pozy
skuje ją głównie z roślin, a w dodatku jego organizm jest zaprogramowany na
bardzo oszczędną gospodarkę wodną. Tymczasem w owym ogrodzie zoologicz
nym podawano gazelom wodę obficie. W rezultacie mocz gerenuka stał się na
tyle rozcieńczony, że osłabił się zapach feromonów (Cherfas, 1984). Jak wia
domo, te specyficzne substancje chemiczne odgrywają ogromną, bodźcową rolę
we wczesnej fazie kojarzenia. Poprzez wąchanie moczu samicy, samiec wie,
w jakiej fazie cyklu rozrodczego znajduje się jego potencjalna partnerka.
W wyniku nieuwzględnienia wiedzy o trybie życia i zachowaniu się gerenuka
nastąpiło przerwanie całego łańcucha sekwencji instynktowego behawioru roz
rodczego, prowadzącego w efekcie do kopulacji.
5. Cyrk
W trakcie pokazów zwierząt w cyrku treser wykorzystuje nie tylko techniki
behawiorystyczne (zob. rozdz. 9), ale także mechanizmy zachowania natural
nego, którym interesuje się etologia.

132
W przypadku pokazu dużych kotów, zwierząt groźnych i w znacznym
stopniu nieprzewidywalnych, treser musi podjąć pewne środki ostrożności.
Jednym z nich jest fakt, iż wchodzi on zawsze jako pierwszy na arenę. Ma to
ogromne znaczenie, ponieważ treser występuje tym samym wobec pojawiają
cych się później kotów jako właściciel terytorium. Wiadomo zaś, że obecność
na obcym terytorium zmniejsza pewność siebie zwierzęcia. Lwy i tygrysy, które
wychodzą na arenę, są więc z góry niejako bardziej uległe wobec człowieka.
W tresurze kotów w sposób częściowo nieświadomy manipuluje się też ich
dystansem ucieczki i krytycznym (zob. rozdz. 3). Treser to zbliża się, to oddala
od zwierząt, wywołując tym samym pożądany przez siebie kierunek ruchu
zwierząt i ich zatrzymywanie się we właściwych miejscach.
6. Laboratorium
We współczesnych laboratoriach niektóre zwierzęta wykorzystuje się do
celów biomedycznych. W doświadczeniach służą one jako tzw. modele (można
na nich obserwować przebieg jakiejś ludzkiej choroby) lub też do testowania
efektów rozmaitych zmian wprowadzonych przez eksperymentatora (np. nowa
torski zabieg leczniczy, zastosowanie nowego leku itd.). Zrozumiałe, że w ocenie
wyników wszystkich tych badaniach dużą rolę odgrywa zachowanie się zwierząt.
Jako przykład wykorzystania zwierząt w badaniach biomedycznych, przy
wykorzystaniu obserwacji ich zachowania się, można przytoczyć eksperymenty
nad LSD. Jest to bardzo silny środek halucynogenny, który z jednej strony bywa
wykorzystywany nielegalnie jako narkotyk, z drugiej - powodując zmiany
w psychice i zachowaniu, podobne jak w schizofrenii, czym wzbudził zaintere
sowanie środowiska medycznego.
W laboratoriach firmy Sandoz przeprowadzono rozległe badania nad wpły
wem LSD na behawior różnych zwierząt - od bezkręgowców do ssaków
(Hoffmann, 1980). Jak można się było domyśleć, efekty narkotyku były różne
i uzależnione od stopnia rozwoju układu nerwowego. U ryb akwariowych
obserwowano więc jedynie nietypową pozycję przy pływaniu, u pająków (przy
większych dawkach) - zaburzenie budowy sieci, natomiast u myszy - kłopoty
w poruszaniu się. Zupełnie inaczej wyglądało zachowanie się zwierzęcia obda
rzonego stosunkowo dobrze rozwiniętą korą mózgową - kota domowego.
Badacze odnotowali jeżenie sierści i ślinienie się, lękliwe wpatrywanie się
przed siebie, brak zainteresowania siedzącą obok myszą lub wręcz lęk przed
nią. Wszystko to wskazywało na występujący u zwierzęcia efekt halucynogen
ny preparatu LSD. Podobnie zachowywał się pies domowy. Warto zwrócić
uwagę, że w tych przypadkach, patrząc z punktu widzenia etologii, pojawił się
133
u zwierzęcia wzorzec zachowania typowy dla strachu, choć w niewłaściwym
kontekście.
Jako zwierzęta laboratoryjne wykorzystywane są głównie: albinotyczne
i barwne odmiany myszy domowej i szczura wędrownego, a także królik, kot
i pies. Jednak także inne gatunki okazują się cenne, przede wszystkim z uwagi
na specyficzny tryb życia i szczególne cechy biologii, które mogą pomóc
w rozwiązaniu problemów medycyny ludzkiej. Niektóre spośród tych gatun
ków wymieniono w tabeli 15. Z dużej grupy zostały wyselekcjonowane jedynie
te zwierzęta, nad którymi prowadzi się badania wyraźnie wiążące się z beha-
wiorem.
Tabela 15. Rzadziej spotykane gatunki zwierząt laboratoryjnych, nad którymi prowadzi się badania
mające związek z behawiorem ( w g różnych źródeł)
Pozycja systematyczna
- gatunek
Miejce
występowania
Pole wykorzystania
1
2
3
Ssaki - naczelne
Sajmiri
(Saimiri spp.)
Ameryka Płd.
model do badania uzależnień (od narkotyków, leków itd.)
Kapucynka czubata
(Cebus apella)
Ameryka Płd.
badanie afiliacji społecznej
Ponocnica
(Aotus spp.)
Ameryka Płd.
badania z zakresu biorobotyki
Makak lapunder
(Macaca nemestrina)
Azja
model do badania choroby A I D S , zmiany występujące
w czasie starzenia się, model do różnorodnych badań
behawioru
Szympans
(Pan troglodytes)
Afryka
badania kognitywne, inteligencja, społeczne
Ssaki - g r y z o n i e
Suwak mongolski
(Meriones
ungidculatus)
- popularny gerbil
Azja
model do badania apopleksji (u niektórych s z c z e p ó w
wsobnych do 1 0 0 % osobników)
Nowiki
(Neotoma spp.)
Ameryka Płn.
preferencje pokarmowe, zachowanie kopulacyjne
Ryżak
(Oryzomys paluslńs)
Ameryka Płn.
zachowanie kopulacyjne
Bawełniak
(Sigmodon hispidus)
Ameryka Płn.
w badaniach różnych rodzajów zachowania
Piżmak
(Ondalra zibethicus)
Ameryka Płn.
Europa
odruch pływania nie zablokowany przez anestezję

134
Tabela 15, cd.
1
2
3
Szynszyla
(Chinchilla laniger)
Ameryka Płd.
zmiany w y w o ł a n e starzeniem się, badania słuchu i jako
model adaptacji do życia na dużych wysokościach
D e g ù
A m e r y k a
Płd.
zachowanie rozrodcze, zabawa
(Octodon degù)
Suseł lamparci
Ameryka Płn.
hibernacja
(Spermophilus
tridecemlineatus)
Pręgowiec amerykański Ameryka Płn
hibernacja
(Tamias striatus)
Ś w i s z c z
Ameryka Płn.
hibernacja
(Marmota monax)
Szczuroskoczki
Ameryka Płn.
model psychotropowego działania leków
(Dipodomys spp.)
Liomysy
(Liomys spp.)
Ameryka Płn.
badanie wpływu lotu kosmicznego na organizm (który
nie musi pobierać wody)
Koszatka
Eurazja
hibernacja
(Dryomys nitedula)
Ssaki - góralki
Góralek abisyński
Afryka
zachowanie pokarmowe przy ograniczonym pobieraniu
(Procavia capensis)
w o d y
Ssaki - m i ę s o ż e r n e
Norka amerykańska
(Mustela visori)
Ameryka Płn.
model dziedzicznej głuchoty u fermowej odmiany białej
Hedlunda
Ssaki - p a r z y s t o k o p y t n e
Dujker modry
(Cephalophus
Afryka
intensywność i efektywność żerowania ssaków przeżu
wających
monitocola)
Ssaki - nietoperze
Mroczek
północnoamerykański
Ameryka Płn.
hibernacja, echolokacja
(Eptesicus fuscus)
Wampir
Ameryka Płd.
zachowanie społeczne
(Desmodus rotundus)
Ptaki
Gołąb d o m o w y
Europa
badanie zdolności orientacji (homing)
(Columba livid)
M i ę c z a k i - g ł o w o n o g i
Ośmiornica
rozpowszech
badania neurologiczne nad rozróżnianiem bodźców
(Octopus binoculariś) niona w akwe
nach świata
wzrokowych i dotykowych

135
7. Dom i gospodarstwo rolne
Zwierzęta te są efektem udomowienia, procesu, w którym zwierzęta dzikie
były brane do gospodarstwa ludzkiego i selekcjonowane na cechę braku agre
sywności i lękliwości względem człowieka. W udomowieniu wiedza o zacho
waniu się odegrała więc podstawową rolę (Belayev, 1974).
Zwierzęta domowe można podzielić na dwie grupy - towarzyszące (ang.
pet animals)
i gospodarskie (ang. farm animals). W pierwszej grupie najważ
niejszym gatunkiem jest pies domowy, który jest efektem udomowienia wilka
i prawdopodobnie w ogóle pierwszym zwierzęciem, które znalazło się w gospo
darstwie człowieka.
Cały behawior psa, który człowiek wykorzystuje w różnych celach, jest
w istocie modyfikacją instynktowego zachowania się wilka. I tak np. stróżowa
nie (pilnowanie terenu, samochodu itd.) jest pochodną zachowania terytorial
nego, kierowanie grupą owiec i pomoc myśliwemu to z kolei modyfikacja róż
nych faz zachowania łowieckiego itd. Żyjąc w rodzinie ludzkiej, pies przenosi
na nią pewne mechanizmy życia watahy wilków, np. wykazuje wobec domow
ników zachowania hierarchiczne przypominające zachowanie swego dzikiego
przodka. Pies wiąże się też z człowiekiem bardzo silną więzią, która także była
utrwalana przez selekcję, a za podstawę więź ta ma mechanizmy afiliacji
w grupie wilków, zwłaszcza relacje osobników podporządkowanych względem
alfa-dominanta.
Także inne zwierzęta towarzyszące (kot, gryzonie, królik, ptaki ozdobne itd.)
wchodzą z człowiekiem w złożone interakcje, które ogólnie można określić
mianem więzi. Według Amerykańskiego Stowarzyszenia Medycyny Weteryna
ryjnej związek ten można zdefiniować jako „wzajemnie korzystną i dynamiczną
relację między człowiekiem a zwierzęciem, na którą wpływ ma zachowanie się
i która jest niezbędna dla zdrowia i dobrego samopoczucia obu stron. Należą do
niej (ale nie ogranicza się) interakcje emocjonalne, psychologiczne i fizyczne
między ludźmi, zwierzętami i środowiskiem". Nawiązując do tej definicji, można
stwierdzić, że kontakt ze zwierzęciem poprawia samopoczucie psychiczne,
a czasem nawet stan zdrowia człowieka, na co wskazują testy kliniczne, np. obni
żone ciśnienie krwi (Robinson - red., 1995).
Także zachowanie się zwierząt gospodarskich od kilkudziesięciu lat budzi
zainteresowanie badaczy. Na początku należy stwierdzić, że każdy gatunek ma
swoją specyfikę, wynikającą z charakterystyki biologicznej przodka, od którego
pochodzi. I tak np. świnia domowa wykazuje, śladem swego przodka - dzika,
intensywne zachowania eksploracyjne, cechę typową dla zwierząt wszystko-
żemych. Ogromną rolę w zachowaniu się koni odgrywa ruch, bo też dzikie
konie mogą często przemierzać znaczne przestrzenie. Natomiast przeżuwacze

136
(owca i bydło) mogą wykazywać rytm aktywności typowy dla dzikich gatunków
kopytnych o takim samym sposobie przyswajania pokarmu (tj. przeżuwaniu).
Ze znacznej liczby przykładów wykorzystania etologii w chowie zwierząt
gospodarskich wybrano tylko niektóre, uznane za najbardziej reprezentatywne
(za Nowicki, 1978):
• Po zachowaniu się owiec można rozpoznać stopień wykorzystania pastwi
ska. Nawet na pastwisku o bardzo dobrej jakości owce pasą się w zwartej
grupie. W miarę jak ilość pokarmu maleje, grupa rozbija się na mniejsze
grupki, a w końcu widać odseparowane od siebie, pojedyncze sztuki.
• Poprzez odpowiednią manipulację można zredukować agresywność
u trzody chlewnej. Świnie są zwierzętami społecznymi, dobrze rozpoznają
cymi osobniki znane i obce. Łączenie osobników pochodzących z różnych
grup prowadzi do wzrostu agresywności i urazów, co może być nasilone
przez uwidaczniające się też u świń zachowania terytorialne. Destrukcyjne
zachowania zwierząt można ograniczyć, wprowadzając osobniki z różnych
grup do nieznanego im wszystkim kojca.
• Zastosowanie bodźców-biostymulantów w czasie krycia lub pobierania
nasienia. Jak stwierdzono w rozdziale 3, w czasie kojarzenia u zwierząt
ogromną rolę odgrywają wszelkiego rodzaju bodźce (węchowe, wzrokowe,
wizualne), których nadawcami są samiec bądź samica. Bodźce pochodzące
od samca mogą przyspieszyć występowanie rui. Te swoiste biostymulanty
wykorzystuje się w chowie świń, odtwarzając z magnetofonu specyficzne
odgłosy wydawane przez knura lub rozpylając zsyntetyzowany, wydzielany
przez jego organizm feromon. Z kolei przy pobieraniu nasienia od buhaja
czy indora proces ten zostaje przyspieszony, gdy samiec widzi znajdującą
się niedaleko samicę.
• Zachowanie się samicy świadczy o tym, czy będzie ona dobrą matką.
Przypadki agresywności wobec własnego potomstwa występują przede
wszystkim u owcy i trzody chlewnej (w tym ostatnim przypadku nawet do
dzieciobójstwa). W obydwu wypadkach, gdy ujawnia się zachowanie pato
logiczne, samice z reguły przyjmują pozycje uniemożliwiające młodym
ssanie. U samicy owcy będzie to stałe ustawianie się frontem wobec jagnię
cia próbującego się zbliżyć do jej boku, natomiast u maciory - leżenie na
brzuchu.
Zachowanie się zwierząt gospodarskich brane jest też pod uwagę w próbie
określenia ich dobrostanu (ang. welfare) w warunkach chowu wielkostadnego.
Przebywające w sztucznych i często drastycznych pod względem nasilenia
bodźców stresujących warunkach zwierzęta domowe wykazują wiele anomalii
behawioralnych, które w mniejszym lub większym stopniu mają wpływ na

137
zdrowotność organizmów i wyniki produkcyjne chowu. Definicja dobrostanu
nie jest prosta, jednak najogólniej pojęcie to zakłada dobre samopoczucie osob
nika i jego zdolność do radzenia sobie z warunkami środowiska. Wydaje się
więc, że oprócz wskaźników hormonalnych i neurologicznych, właśnie beha-
wior zwierzęcia jest źródłem informacji o całokształcie jego interakcji ze śro
dowiskiem.
8. Człowiek
Biologicznymi aspektami zachowania się człowieka zajmuje się relatywnie
nowa dyscyplina wiedzy - etologia człowieka (albo humanetologia). W analizie
ludzkiego behawioru traktuje ona nasz gatunek jako jeden z wielu, które wyewo
luowały na Ziemi, stąd też interesuje się porównaniem zachowania ludzkiego
w stosunku do bliższych i dalszych zwierzęcych krewniaków człowieka (z grupy
naczelnych i innych ssaków).
Ulubionym materiałem badań dla etologii człowieka jest niemowlę, u któ
rego można zaobserwować wiele wzorców behawioru w stanie niejako „biolo
gicznie czystym", jeszcze nie przytłumionym lub zmodyfikowanym przez
środowisko (społeczeństwo, kulturę, w których się będzie wychowywać). Jed
nym z najbardziej ważkich dokonań w tej dziedzinie wiedzy jest odkrycie, iż
człowiek przychodzi na świat z wrodzoną zdolnością do identycznego wyrażania
emocji za pomocą mimiki, nawet wtedy, gdy rodzi się jako niewidomy i nieza
leżnie od kultury, w jakiej się wychowuje (Eibl-Eibesfeld, 1984).
W zachowaniu noworodka ludzkiego zastanawia też wiele odruchów (określanych jako atawi- i
styczne lub archaiczne), które zanikają w dalszych fazach rozwojowych dziecka. Tak np. gdy p o d - 1
trzymywany dotyka nóżkami jakiejś płaskiej powierzchni, noworodek wykonuje takie ruchy, jak I
przy chodzeniu. Potrafi on także zaskakująco mocno uchwycić podsunięty mu palec lub inny podobny
obiekt. Siła chwytu jest tak duża, że noworodek może zawisnąć w powietrzu, trzymając się rączkami
linki. Etolodzy są skłonni interpretować takie odruchy atawistyczne jako d o w ó d pokrewieństwa
człowieka z innymi gatunkami naczelnych. Zwracają uwagę, że małpięta po urodzeniu, dzięki
swym chwytnym umiejętnościom, pozostają przez długi czas „przytwierdzone" do piersi lub grzbietu
matki. Badacze o innej orientacji natomiast utrzymują, że odruchy atawistyczne sąjakby przygoto
waniem do normalnej aktywności ruchowej w okresie dojrzałości (Mehler i Dupoux, 1990).

Inne szkoły badań
zachowania się
zwierząt
9.1. Ekologia behawioralna
Ekologia ta powstała w dużej mierze jako wynik inspiracji niektórymi pyta
niami, które stawiali sobie etolodzy w okresie „klasycznym".
Kładzie ona nacisk na aspekt funkcjonalny zachowania (jaką korzyść powo
duje ono dla osobnika?) i postrzega je w perspektywie ewolucyjnej, a jednocze
śnie stara się zinterpretować behawior jako odpowiedź na wymogi stawiane
przez dane środowisko. Innymi słowy, jest to odpowiedź na pytanie: J a k okre
ślony wzorzec zachowania przyczynia się do przeżycia i do sukcesu reproduk
cyjnego danego osobnika?" (Krebs i Davies, 1993; tł. M. Golachowski).
Ekolodzy behawioralni badają porównawczo strategie zachowania się spo
krewnionych gatunków zwierząt w różnych środowiskach. Wspomniany już
wcześniej (rozdz. 3) wpływ rodzaju pokarmu na życie społeczne u naczelnych
jest właśnie przykładem takiego studium.
Ekologia behawioralna bada również procesy optymalizacji behawioru.
Wynika ono z założenia, że zachowując się w określony sposób, zwierzę zyskuje
coś, ale również ponosi koszty. Wybór określonej strategii w trakcie np. zdoby
wania pokarmu czy partnera jest więc podyktowany bilansem zysków i kosz
tów („decyzją ekonomiczną"). Nie oznacza to jednak, że zwierzę świadomie
dokonuje takiego bilansu. Należy raczej powiedzieć, że każde zachowanie się
ma przynależne sobie pewne zyski i koszty, natomiast wybór, jaki dokonuje się
między alternatywnymi rodzajami behawioru, jest wynikiem doboru naturalnego.
Wiele przykładów strategii optymalizacji opisano w przypadku zdobywania
pokarmu. Jeden z najbardziej interesujących dotyczy żerowania wyjca płaszczo
wego (Alouatta paliata) w lasach Kostaryki. Małpa ta, mimo iż żywi się liśćmi,
wykazuje ogromną wybiórczość wobec potencjalnego pokarmu. Wyjca, po pro
stu, otaczają liście, a mimo to wybiera tylko niektóre drzewa, a z tych drzew -
jedynie niektóre listki (Glander za Alcock, 1993). Taki sposób żerowania jest
czaso- i energochłonny (koszt), a jednak tylko tym sposobem zwierzę może
uniknąć substancji toksycznych, przede wszystkim alkaloidów, utrudniających

139
przyswajanie białka z pokarmu (zysk). Jeśli zważyć, że chore czy osłabione
zwierzę w naturalnym środowisku szybko ginie, preferowanie tej właśnie stra
tegii staje się oczywiste.
Jednak nie zawsze mamy do czynienia z tak klarowną sytuacją, na co wska
zuje przykład zachowania się ostrygojada - Haemoptopus ostralegus. Ptak ten
żeruje na wybrzeżu i żywi się (jak wskazuje nazwa) mięczakami, których
muszle otwiera uderzeniami dzioba albo uderzeniami o ziemię. W skuteczności
tego zachowania najistotniejsze jest, jak wielką (pod względem wymiarów)
zdobycz wybiera ptak. Największe mięczaki wymagają więcej czasu, by otwo
rzyć muszle (koszt), za to stanowią pokarm bardziej kaloryczny (zysk). Na pod
stawie przeprowadzonej kalkulacji wydawało się więc, że wybór największego
pokarmu to właściwa strategia optymalizacji. Jednak hipoteza ta nie została zwe
ryfikowana pozytywnie. Okazało się bowiem, że ostrygojad zjada mięczaki
w sumie nieco większe niż przeciętne, ale nie największe. Na takie zachowanie się
miały wpływ czynniki nie brane pod uwagę w założeniach wstępnych: (1) ptak
czasem wybierał największe mięczaki, ale zachowywał się „ekonomicznie" i gdy
nie mógł ich otworzyć dość szybko, rezygnował i poszukiwał innych, (2) im
większa muszla, tym większe było prawdopodobieństwo, że znajdują się na niej
pąkle (Cirripaedia) niejadalne dla ostrygojada, a utrudniające otwieranie muszli
(Meire i Envynck, 1986).
Warto wspomnieć też o zyskach i kosztach związanych z zachowaniem
społecznym, ze względu na szeroko dyskutowane zjawisko altruizmu. Z punktu
widzenia „decyzji ekonomicznej" jest to specyficzne zachowanie się, w którym
jeden osobnik ponosi koszty, by inny zyskiwał (Buss, 1999).
Przyrodnikom od wielu lat znane były zachowania wielu gatunków, u któ
rych osobniki określonej grupy czy społeczności angażowały się w aktywność,
która mogła im przynieść śmierć, a zysk - innym (i nie był to behawior rodzi
cielski). Ostrzeganie o zbliżającym się drapieżniku, które może spowodować
zabicie przez drapieżnika osobnika ostrzegającego (niektóre gryzonie żyjące
w koloniach) czy angażowanie się w ryzykowne dla każdego z łowców polo
wania grupowe (lwy) to tylko niektóre przykłady altruizmu. Początkowo dość
łatwo interpretowano je jako zachowania służące przetrwaniu gatunku. Jednak
teoretycy ewolucjonizmu i genetyki populacji wykazali, że tak zwane przetrwa
nie gatunku jest fikcją, gdyż selekcja naturalna działa na poziomie osobniczym,
a zatem to każdy osobnik stara się przeżyć i przekazać następnym pokoleniom
własne geny. Tym samym zachowania altruistyczne stały się prawdziwą zagadką
(Alcock, 1993).
Przełom przyniosło opracowanie przez W.D. Hamiltona teorii dostosowania
łącznego.
Najkrócej mówiąc, mówi ona, że określone zwierzę ma dwie drogi
przekazywania swoich genów do następnych pokoleń - reprodukcję własną

140
albo pomoc w przetrwaniu osobnikom, które choćby w części są nosicielami
tych samych, co on, genów. Te inne osobniki to oczywiście rozmaici krewniacy
(Buss, 1999).
Teoretyczne założenia altruizmu zostały potwierdzone między innymi
dzięki badaniom nad żyjącym grupowo gryzoniem z rodziny wiewiórkowatych
- susłem Beldinga (Spermophilus beldingi). Na widok drapieżnika „naziemnego"
(jak kojot czy borsuk) osobnik, który zauważył niebezpieczeństwo, wydaje
specyficzny okrzyk - jest on dla innych członków grupy sygnałem do ucieczki.
Zatem szansa przeżycia innych susłów wzrasta, ale osobnik podnoszący alarm
naraża się na atak drapieżnika i być może na śmierć. W wyniku analizy obserwa
cji polowych okazało się, że susłami wydającymi sygnał alarmowy są znacznie
częściej samice, których więź z własnymi krewniakami jest bardzo mocna
(związek samców z resztą grupy jest natomiast luźny). Choć alarmując, samica
naraża własne życie, jednak pomaga przeżyć osobnikom spokrewnionym, które
w pewnej mierze są nosicielami jej genów (Shernan, 1977).
Podane wyżej przykłady wyjaśnienia zachowań dokonywane przez ekolo
gów behawioralnych są jedynie niektórymi wybranymi spośród wielu dokonań
tej młodej, ale bardzo obiecującej dziedziny wiedzy.
9.2. Behawioryzm
Jest to nurt, którego początki sięgają badań I. Pawłowa (1849-1936) w Rosji
(a później w ZSRR) i J. Watsona (1878-1958) w USA - skoncentrował się na
zagadnieniu uczenia się. Podstawą badawczą klasycznego behawioryzmu są
obserwacje wyzwalanego przez kontrolowane bodźce zachowania się, nato
miast rola towarzyszących stanów wewnętrznych zwierząt i ludzi (np. emocji)
jest marginalizowana. Behawioryzm eksponuje wpływ środowiska na beha-
wior, natomiast minimalizuje znaczenie genów. W dużym stopniu negowana
jest także różnica w zachowaniu, wynikająca z mniej czy bardziej zaawanso
wanej budowy układu nerwowego. Wyniki badań przeprowadzone na szczu
rach laboratoryjnych można więc, według niektórych badaczy tego nurtu,
uogólnić na człowieka.
Badania behawiorystów były i są prowadzone w laboratoriach, a ich przed
miot to właśnie typowe zwierzęta laboratoryjne (gryzonie, pies, kot, gołąb i inne).
W nurcie tym historycznie można wyróżnić dwie szkoły - warunkowanie kla
syczne (ang. classical lub pavlovian conditioning) oraz warunkowanie instru-

141
mentalne (ang. operant conditioning). Wyniki badań obu szkół są szeroko wyko
rzystywane w tresurze zwierząt.
Warunkowanie klasyczne. Opiera się ono na ukształtowaniu reakcji zwie
rzęcia według asocjacji (kojarzenia) bodźców. Każdy organizm przejawia
szereg prostych reakcji na bodźce, które są zaprogramowane genetycznie. Są to
tzw. odruchy bezwarunkowe. U ssaków to np. ślinienie się na widok pokarmu,
zmniejszenie źrenicy pod wpływem silnego światła, prostowanie kończyny
przy uderzeniu w rzepkę kolanową itd.
Jeśli bodźcowi, który powoduje reakcję bezwarunkową, towarzyszy jakiś
inny, neutralny bodziec (np. dzwonek), to po określonej liczbie powtórzeń
wywołania reakcji następuje skojarzenie obu tych bodźców.
W efekcie, po pewnym czasie już sam bodziec, do tej pory nie znaczący
(czyli w tym przykładzie dzwonek), wywołuje reakcję. Tym samym kształtuje
się odruch warunkowy.
W klasycznym doświadczeniu I. Pawłowa, kiedy pies dostawał mięso, ślinił
się. Kiedy podawaniu mięsa towarzyszył dzwonek i gdy procedurę tę powta
rzano określoną liczbę razy, pies w końcu ślinił się na dźwięk dzwonka, choć
nie dostawał mięsa. U zwierzęcia wytworzyła się zatem zdolność do reagowa
nia w sposób odruchowy w odpowiedzi na bodźce, które z określonym zacho
waniem normalnie nie mają nic wspólnego.
Kształtowanie się odruchu warunkowego w wariancie klasycznym można
przedstawić na podstawie poniższego schematu, w którym podano także stoso
waną w behawioryzmie terminologię:
1. Odruch bezwarunkowy
Bodziec bezwarunkowy
(inaczej wzmocnienie)
(mięso)
-> Reakcja bezwarunkowa
(ślinienie się)
2. Kształtowanie odruchu warunkowego
Bodziec bezwarunkowy
(mięso)
+ • Reakcja bezwarunkowa
Bodziec obojętny
(inaczej warunkowy)
(dzwonek)
(ślinienie się)
3. Odruch warunkowy
Bodziec warunkowy • Reakcja warunkowa
(dzwonek)
(ślinienie się)

142
Badając procesy uczenia metodą warunkowania, Pawłów wykrył wiele pra
widłowości. Na przykład, by efektywnie ukształtować odruch warunkowy,
bodziec warunkowy powinien (w czasie) poprzedzać bodziec bezwarunkowy
albo oba te bodźce powinny przebiegać równocześnie. Podobnie uczony ten
wskazał na bardzo istotną rolę wzmocnienia (czyli bodźca bezwarunkowego).
Zostało potwierdzone, że jeśli reakcji warunkowej nie towarzyszy od czasu do
czasu wzmocnienie, ma ona tendencję do zaniku, czyli wygaszania. W omawia
nym wyżej przykładzie oznacza to, że reakcji ślinienia się na dzwonek powinno
towarzyszyć, co pewien czas, podawanie mięsa. Interesującym odkryciem było
również stwierdzenie faktu, że reakcja bezwarunkowa pod względem ilościo
wym nie jest równa warunkowej - objętość wydzielanej przez psa śliny w obu
przypadkach była różna.
Obecnie istnieją dwie teorie tłumaczące, w jaki sposób kształtuje się skojarzenie między bodź
cem warunkowym i bezwarunkowym (Forkman, 2002). Pierwsza z nich głosi, że bodziec warun
k o w y powoduje pojawienie się w mózgu zwierzęcia obrazu nagrody, a to z kolei wywołuje reakcję
warunkową. Tak więc, by odwołać się do p o w y ż s z e g o przykładu, tworzy się ciąg: dzwonek-obraz
mięsa-ślinienie się. Według alternatywnego poglądu bodziec warunkowy staje się odpowiednikiem
bezwarunkowego. Tak więc dzwonek jak gdyby przejmuje cechy mięsa i wyzwala reakcję ślinienia
się. W pierwszym przypadku można więc powiedzieć, iż zwierzę w jakiś sposób „wie", że dzwonek
zapowiada pokarm, w drugim - po prostu reaguje na obecność dzwonka bardzo podobnie, jak na
uprzednie podawanie mięsa. W świetle wielu przeprowadzonych do tej pory doświadczeń wydaje
się, że u zwierząt pojawia się raczej pierwszy typ skojarzenia.
Dalsze badania przedstawicieli „szkoły pawłowowskiej" wykazały, że ten
rodzaj warunkowania może występować zarówno u tak nieskomplikowanych
pod względem morfologii zwierząt, jak płazińce, jak i u człowieka. Proces warun
kowania klasycznego można także wywołać w czasie ciąży u płodu ludzkiego
(Hilgard i Marquis, 1961).
Warunkowanie instrumentalne. Oprócz wspomnianego wyżej J. Watsona,
do rozwoju szkoły warunkowania instrumentalnego przyczynili się tacy bada
cze amerykańscy, jak: E. Thorndike (1874-1949), E. Tolman (1886-1959)
i B. Skinner (1903-1990). W USA zostały też opracowane oryginalne procedury
badawcze, polegające na prowokowaniu zachowania się zwierzęcia, a ściślej
rozwiązywania określonego problemu (odnalezienie drogi do pokarmu w specjal
nym labiryncie, naciśnięcie dźwigni dla uzyskania pokarmu itd.; rys. 12).
Ujmując najprościej, warunkowanie instrumentalne jest rodzajem uczenia,
w którym behawior zmienia się poprzez swoje konsekwencje. Kiedy zwierzę
zachowuje się w określony sposób, skutki tego oddziaływają tak, że zachowa
nie utrwala się albo zanika. Podczas gdy warunkowanie klasyczne polegało na
kojarzeniu bodźców, to naczelną zasadą warunkowania instrumentalnego jest
asocjacja między behawiorem a jego skutkiem.
143
dystrybutor pokarmu dźwignia
(granulat) głośnik
karmi dto
podłoga rusztowa z możliwością
podłączenia do prądu elektrycznego
Rysunek 12. Typowe doświadczenia stosowane w badaniach warunkowania instrumentalnego:
a - labirynty - zwierzę musi opanować najkrótszą drogę prowadzącą do pokarmu, b - skrzynka
Skinnera - zwierzę zostaje wynagrodzone lub ukarane w konsekwencji określonego zachowania
(naciśnięcie dźwigni, pozostawanie w miejscu narażonym na podrażnienie przez prąd elektryczny)
(wg różnych źródeł)

144
W klasycznych eksperymentach prowadzonych przez behawiorystów można
zobaczyć, że zwierzę przyswaja sobie tę relację stopniowo, a reakcja kształtuje
się poprzez próby i błędy. Na przykład, szczur w skrzynce problemowej (rys. 12)
początkowo porusza się w sposób chaotyczny aż do momentu, gdy przypadkowo
naciśnie dźwignię, co spowoduje automatyczne podanie pokarmu. Prowadzi to
w końcu do celowego działania zwierzęcia - naciskania dźwigni w celu uzyska
nia pokarmu.
W innych przypadkach zwierzę zostaje stopniowo naprowadzane na czyn
ność, którą musi wykonać. Przy warunkowaniu instrumentalnym zwierzę powin
no być motywowane (np. głodne), tylko wtedy bowiem można je skłonić do
wykonania zadania.
Gdy skutek zachowania się jest pozytywny dla zwierzęcia - mówi się
o wzmocnieniu dodatnim albo nagrodzie. Nagroda (w doświadczeniach z warun
kowaniem - najczęściej pokarm), którą uzyskuje osobnik po wykonaniu jakiejś
czynności, powoduje, że zwierzę będzie tę czynność wykonywać chętnie i jest
duże prawdopodobieństwo, że częstotliwość jej występowania wzrośnie. I prze
ciwnie, skutek zachowania się może być dla zwierzęcia negatywny (np. podraż
nienie prądem elektrycznym). Jest to kara. Wydaje się wielce prawdopodobne,
że behawior karany będzie słabł, a jego częstotliwość malała.
Zarówno nagradzanie, jak i karanie może następować nie tylko przez zadzia
łanie jakimś bodźcem, lecz przez odjęcie bodźca już działającego.
Tak więc istnieją teoretycznie cztery możliwości nagradzania i karania,
które zostały przedstawione w poniższym schemacie, łącznie z przykładami
zaczerpniętymi z tresury psa domowego:
Element zadania
Nagroda
(powoduje zwiększenie
częstotliwości behawioru)
Kara
(powoduje zmniejszenie
częstotliwości behawioru)
Zostaje wprowadzony
wprowadzony element powoduje
zwiększenie częstotliwości beha
wioru;
podawanie przysmaku za wykona
nie zadania powoduje utrwalenie
odpowiedniego zachowania się
wprowadzony element powoduje
zmniejszenie częstotliwości zacho
wania;
nagana powoduje zaniknięcie niepo
żądanego zachowania
Zostaje odjęty
odjęty element powoduje zwiększe
nie częstotliwości behawioru;
ulga czy poluzowanie może działać
wzmacniająco na behawior,
pies z poluzowaną smyczą chętnie
idzie obok właściciela
odjęty element powoduje zmniej
szenie częstotliwości zachowania;
większe zdyscyplinowanie działa
osłabiająco na niepożądany beha
wior;
pies wzięty na smycz po akcie niepo
słuszeństwa

145
Gdy za pomocą warunkowania instrumentalnego u zwierzęcia ukształtuje
się już pożądane zachowanie, procedurę można skomplikować w ten sposób, że
zwierzę nie jest nagradzane po każdej próbie. Mówi się wtedy o tzw. wzmocnie
niu częściowym. Wracając do powyższego przykładu, dla uzyskania pokarmu
szczur będzie ponawiał próbę naciskania dźwigni, ale nagrodę (w zależności od
rodzaju procedury) dostanie już tylko wtedy, gdy czynność tę wykona określoną
liczbę razy albo co pewien czas (np. w interwałach 10-minutowych). Te nowe
zasady musi opanować, by nie podejmować stałych i bezowocnych wysiłków.
W praktyce techniki warunkowania klasycznego i instrumentalnego często
łączy się ze sobą. Dzieje się tak np. wtedy, gdy w trakcie nauki zwraca się zwie
rzętom uwagę na pożądane zachowanie się. Jeśli np. treser pracuje z delfinem,
znajduje się on w pewnej odległości od zwierzęcia i musi dać sygnał, że okre
ślone zadanie zostało wykonane prawidłowo i zwierzę powinno przybyć po
odebranie nagrody. Sygnałem takim będzie np. dźwięk gwizdka. Jednak by
zwierzę zaczęło zwracać uwagę na ten bodziec, należy go tego najpierw nauczyć
metodą warunkowania klasycznego. Trzeba więc wytworzyć skojarzenia
dźwięku gwizdka z przybyciem po nagrodę, posługując się techniką taką, jaką
zastosowano w doświadczeniu z wydzielaniem się śliny u psa.
Na skuteczność obu typów warunkowania wpływa kilka czynników: czas,
który upływa między działaniem bodźca warunkowego i bezwarunkowego
(warunkowanie klasyczne), albo między reakcją a nagrodą lub karą (warunkowa
nie instrumentalne), siła wzmocnienia i specyficzne cechy bodźca warunkowego
oraz predyspozycje zwierząt do kojarzenia określonych bodźców. Szczególnie
warta podkreślenia jest ta ostatnia cecha. Badania na szczurach laboratoryjnych
wykazały np., że zwierzęta te łatwo kojarzą ze sobą dźwięk i ból oraz smak
i wymioty, natomiast nie mogły wytworzyć skojarzenia między dźwiękiem
i wymiotami oraz smakiem i bólem (Garcia i Koelling za Forkman, 2002).
Behawioryzm odegrał ogromną rolę w psychologii, kładąc podwaliny pod
naukowe badania procesów uczenia się zwierząt i ludzi. W sferze praktycznej
pozwolił także na opracowanie różnorodnych technik tresury. Pod tym wzglę
dem na uwagę zasługują osiągnięcia badaczy amerykańskich K. i M. Brelan-
dów. Udowodnili oni, iż metodą warunkowania instrumentalnego można na
uczyć rozmaitych złożonych czynności wiele różnych zwierząt, takich jak kura
domowa, królik czy szop pracz. W trakcie swojej działalności Brelandowie
wytresowali ponad 6000 osobników rozmaitych gatunków zwierząt, wykorzy
stując swój sukces także komercyjnie (pokazy na potrzeby telewizji, w domach
towarowych itd.).

146
Jednak to właśnie Brelandowie zwrócili uwagę na ograniczenie metody
warunkowania i na rolę odziedziczalnych form zachowań w życiu zwierząt.
W tresurze napotykali na trudności i trafnie rozpoznawali ich charakter.
Stwierdzili, że w zachowaniu ich podopiecznych występuje rodzaj regresji do
behawioru instynktowego (Breland i Breland, 1961). Na przykład, u jednego ze
zwierząt, świni, warunkowano reakcję przenoszenia w pysku drewnianych
„monet". Brelandowie zauważyli, że po pewnym czasie świnia ta wykonywała
nagradzaną czynność coraz dłużej, porzucała po drodze przedmiot i zajmowała
się ryciem, które, jak wiadomo, należy do podstawowego repertuaru behawioru
tego zwierzęcia. Wskazuje to na pewne - przynajmniej w wypadku zwierząt -
granice efektów uczenia, które można osiągnąć metodami behawiorystycznymi.
9.3. Kognitywizm
Jest to kierunek łączący analizę procesów przetwarzania informacji (po
strzeganie, myślenie, uczenie się, planowanie itd.) z badaniami mózgu („w jaki
sposób mózg pracuje, tak jak pracuje?") i teorią informacji (np. modelowanie
procesów funkcjonowania umysłu). O zastosowaniu kognitywizmu w bada
niach dotyczących zachowania się zwierząt była mowa w części poświęconej
inteligencji (rozdz. 3).

10.
Obserwacje
zachowania się
zwierząt
10.1. Uwagi metodyczne
Końcowe uwagi nie są przewodnikiem metodycznym w zakresie badań
behawioru zwierząt, lecz jedynie próbą zasygnalizowania specyfiki takiej pracy.
Biorąc natomiast pod uwagę ogromny nacisk, jaki obecnie kładzie się na etyczną
stronę badań naukowych, zaprezentowano poniżej pewne zasady i ograniczenia
badań, płynące właśnie z inspiracji etycznej, a zalecane przez Amerykańskie
Towarzystwo Badań Behawioru Zwierząt (Animal Behaviour Society).
Wybór gatunku
W przypadku zwierząt wiwaryjnych zasada ogólna głosi, że przy rozwiązy
waniu określonego problemu badawczego za obiekt wybiera się raczej gatunki
o dobrze poznanej biologii i sprawdzone w praktyce laboratoryjnej. Natomiast
unikać należy zwierząt egzotycznych, słabiej poznanych i nierzadko trudniej
szych w utrzymaniu. Reguła ta nie dotyczy oczywiście tych gatunków, które ze
względu na swoją biologiczną specyfikę są doskonałymi modelami dla badania
jakiegoś problemu związanego z behawiorem, choć nie zaliczają się do typo
wych zwierząt laboratoryjnych (zob. tab. 15 - rozdz. 8).
Badania behawioru zwierząt w środowisku naturalnym i w ogrodzie zoolo
gicznym służą zwykle poszerzeniu wiedzy o określonym gatunku, a także
o złożonych interakcjach, w które wchodzi (np. zależności pokarmowe). Ograni
czenia obserwacji w naturze mają przede wszystkim charakter etyczny (zwie
rzęta zagrożone wyginięciem, możliwe zakłócenia wywołane przez obserwacje),
o czym będzie mowa w dalszej części rozdziału.
Harmonogram obserwacji
Wstępną fazą obserwacji behawioru określonego gatunku zwierzęcia jest
zawsze zapoznanie się z jego biologią na podstawie dostępnej literatury. Chodzi
o podstawowe dane dotyczące sposobu życia, typu aktywności, wskaźników

148
reprodukcyjnych itd. Jeśli informacje takie są skąpe, lub niedostępne, należy
zapoznać się z danymi literaturowymi, dotyczącymi gatunków pokrewnych.
Pierwszą fazą badań są obserwacje wprowadzające, tzw. pilotowe. Mają one
na celu przede wszystkim określenie pory, w którym zwierzę jest widoczne
(tj. nie chowa się w jakiejś kryjówce) i wykazuje aktywność, jak również przy
bliżenie obserwatorowi specyfiki behawioru danego zwierzęcia. W tym miejscu
warto dodać, że u wielu gatunków występuje znaczna elastyczność i zmienność,
jeśli chodzi o aktywność dobową (mogą np. zmieniać dzienny na nocny tryb
życia) i ujawnianie różnorodności form behawioru. Zmiany takie mogą wystę
pować zarówno w warunkach wiwaryjnych (ogród zoologiczny), jak i nawet
w różnych warunkach środowiska naturalnego.
Technika obserwacji zasadniczej uzależniona jest od projektu badawczego.
Jeśli chce się np. określić budżet czasu i etogram osobnika(ków) określonego
gatunku, zaleca się prowadzenie obserwacji ciągłych i wyrywkowych. W spo
sób ciągły rejestruje się zachowanie aktywnego zwierzęcia w bloku kilkugo
dzinnym (najlepiej 4-godzinnym) w ciągu przynajmniej kolejnych, trzech dni.
Uzyskane wyniki uzupełniane są badaniami wyrywkowymi, w których w spo
sób losowy wybiera się i przeprowadza w kilkugodzinnym bloku pewną licz
bę krótkich (maksimum 5-minutowych) obserwcji. Konfrontacja rezultatów
obserwacji ciągłych i wyrywkowych daje pełniejszy obraz zarówno różno
rodności behawioru, jak i czasu poświęcanego przez zwierzę na poszczególne
czynności.
Technika obserwacji
Regułą ogólną jest znalezienie takiego miejsca do prowadzenia obserwacji,
które umożliwia dobry widok obiektu, a jednocześnie w jak najmniejszym stop
niu wpływa na zachowanie się zwierzęcia. W środowisku naturalnym obserwator
ma często do czynienia z gatunkami czujnymi i płochliwymi. Wymusza to okre
ślony sposób prowadzenia badań, czasem w specjalnie przygotowanej kryjówce
(np. w ambonie - osłoniętej platformie umieszczonej nad ziemią). W innych
wypadkach należy zachowywać inne środki ostrożności (odpowiedni dystans,
zbliżanie się pod wiatr itp.). Można także dodać, że badania terenowe często
wymagają od badacza oprócz przygotowania teoretycznego także specyficz
nych kwalifikacji, jak orientacja w terenie czy odnajdywanie i badanie śladów.
Zwłaszcza ta druga umiejętność jest bardzo pożyteczna. Różnorodne ślady pozo
stawiane przez zwierzęta (tropy, rysy po pazurach na drzewie, miejsca czochra
nia się itd.) mogą bowiem dostarczyć nie mniej informacji o behawiorze niż
bezpośrednie obserwacje.

149
Innego typu problemy można napotkać, prowadząc badania zachowania się
zwierząt w ogrodzie zoologicznym. Wpływ zwiedzającej publiczności na beha-
wior eksponowanych tam gatunków jest często bardzo znaczący. Z jednej strony
efektem tego jest żebranie o pokarm (np. u niedźwiedzi, naczelnych) i inne
anomalie behawioralne, z drugiej - taka zmiana aktywności zwierzęcia, która
umożliwia mu uniknięcie konfrontacji ze zwiedzającymi w godzinach najwięk
szego nasilenia ruchu w zoo (niektóre małe drapieżniki). Harmonogram obser
wacji powinien uwzględnić właśnie te czynniki.
W badaniach behawioru pomocny bywa odpowiedni osprzęt techniczny.
Dobrej klasy urządzenia optyczne (lornetka, luneta), noktowizor, nie mówiąc już
0 urządzeniach do radiotelemetrii, bardzo poszerzają możliwość obserwacji
1 umożliwiają uzyskanie wartościowych wyników. W wielogodzinnych sesjach
badawczych bardzo pomocny jest sprzęt do filmowania techniką wideo. Z kolei
kieszonkowy magnetofon, dyktafon czy rodzaj specjalistycznego palmtopu
umożliwiają rejestrowanie obserwacji bez odrywania oczu od obiektów. Warto
jednak pamiętać, że w niektórych warunkach urządzenia techniczne mogą się
okazać zawodne, natomiast wciąż jeszcze doskonale sprawdza się staroświecki
notatnik i ołówek.
Każdorazowo efektem obserwacji jest spisany protokół. Zawiera on nie
tylko zarejestrowane w danym dniu dane dotyczące behawioru oraz datę, miej
sce i godziny badań, ale także panujące wówczas warunki (jak np. faza Księżyca,
pogoda, a w ogrodach zoologicznych również liczba zwiedzających znajdująca
się w określonych wcześniej momentach obok pomieszczenia zwierzęcia).
Uzyskane dane
Najczęściej można je zaliczyć do trzech grup tematycznych:
• częstotliwość obserwowanych zachowań - ile razy w jednostce czasu
określony osobnik wykazywał dany rodzaj behawioru?
• sekwencji zachowania - jaka była kolejność następujących po sobie
zachowań?
• komunikacji między osobnikami - w jaki sposób każde zwierzę odpo
wiadało na sygnał ze strony wszystkich innych?
Dane opracowuje się statystycznie za pomocą różnych metod (np. analiza
wariancji, testy nieparametryczne). Obecnie dostępne są pakiety programów
komputerowych przeznaczone do analizowania danych behawioralnych, także
tych zarejestrowanych na taśmie wideo.

150
10.2. Kwestie etyczne związane z badaniami
behawioru zwierząt
Nie ulega wątpliwości, że badania zachowania się zwierząt mają swoją
wagę poznawczą i dużą wartość aplikacyjną. Jednocześnie jednak niektóre z nich
mogą budzić wątpliwości natury etycznej. Pojawiają się one wówczas, gdy
do procedury badawczej włączone są elementy manipulacji zwierzętami, np.
schwytanie celem założenia nadajników radiowych, dzięki którym będzie póź
niej można śledzić aktywność zwierząt. W takich wypadkach należy zawsze
ważyć spodziewaną wartość naukową badań w stosunku do możliwego, nieko
rzystnego wpływu procedury badawczej na zwierzę, jego zdrowotność i póź
niejszy rozród, a tym samym szerzej, także na populację gatunku.
Należy pamiętać o pewnych ogólnych zasadach postępowania:
• W badaniach behawioralnych, tak jak w innych, muszą być uwzględnione
wszystkie krajowe akty prawne dotyczące zwierząt (np. ustawy chroniące
prawa zwierząt).
• Przy wyborze gatunku do badań istotny jest jego status zagrożenia i wpis do
międzynarodowej i/lub krajowej „Czerwonej księgi" (rejestr gatunków za
grożonych wyginięciem). Jest jasne, że należy unikać wszelkiego niepoko
jenia przedstawicieli gatunków zagrożonych i wybierać raczej te, które wy
stępują pospolicie.
• Jeśli badania prowadzi się na zwierzęciu utrzymywanym w warunkach
wiwaryjnych, powinno się preferować osobniki urodzone w niewoli, a nie
odłowione z naturalnego środowiska. Warunki utrzymania takich zwierząt
(w klatkach, wolierach itd.) określają regulacje prawne.
• Jeżeli stosuje się procedury badawcze, które mogą wywołać dyskomfort, ból
i stres, powinno się wybrać gatunek najbardziej odporny na te przypa
dłości.
• Chociaż w badaniach behawioralnych przeważają obserwacje na żywych
zwierzętach, w niektórych przypadkach (np. dla celów edukacji) można je
zastąpić, stosując nagrania wideo pochodzące z innych obserwacji lub symu
lacje komputerowe.
• Liczbę osobników uczestniczących w badaniu można ograniczyć dzięki
obserwacjom pilotowym, dobremu zaplanowaniu metodyki i zastosowaniu
procedur statystycznych, które pozwalają przebadać wpływ na zachowanie
się jednocześnie kilku czynników.
• Należy wstępnie określić źródła i charakter możliwego cierpienia badanych
zwierząt.

151
Warto także wymienić niektóre kwestie szczegółowe:
Badania w terenie związane z manipulacjami mogą spowodować ujemne
skutki nie tylko dla pojedynczych zwierząt, ale i dla całej populacji. Chwy
tanie, znakowanie, zakładanie nadajników, pobieranie próbek krwi i inne
tego typu działania wiążą się z ryzykiem późniejszej, mniejszej przeżywal-
ności zwierzęcia i zaburzenia jego rozrodu. W tym wypadku szczególnie
zalecane są badania pilotowe.
Przy odławianiu zwierzęcia z natury do celów obserwacyjnych należy
uwzględnić konsekwencje tego działania dla innych zwierząt (np. dla potom
stwa).
Powinno się unikać wymuszania (poprzez procedurę doświadczalną) inter
akcji między osobnikami tego samego gatunku lub różnych gatunków,
które mogą wiązać się z zadawaniem cierpienia, zranienia czy nawet śmierci
jednego ze zwierząt. W związku z tym drapieżnictwo i zachowania ago-
nistyczne (z walkami na serio) należy badać raczej w środowisku natural
nym, a nie aranżować je na potrzeby badań.
Powinno się ograniczać wszelkiego typu doświadczenia, które wiążą się
z wymuszaniem motywacji poprzez pozbawianie zwierzęcia np. pokarmu
lub działając na nie bodźcami stresującymi.
Powyższa uwaga dotyczy także doświadczeń zaburzających normalne za
chowania społeczne (pozbawianie partnera, izolacja, nadmierne zagęszcze
nie).
Niektóre doświadczenia (np. medyczne) wiążą się z obserwacjami za
chowania zwierząt pod wpływem czynników potencjalnie letalnych (sub
stancje chemiczne, większe dawki leków itd.). W wypadku wyraźnie szko
dliwego i stresującego wpływu czynnika na organizm zwierzęcia zaleca się
stałą obserwację jego stanu zdrowia, leczenie, a w razie konieczności
humanitarne zabicie, możliwie jak najwcześniej, by zredukować cierpienie.
Kwestie te regulują akty prawne dotyczące zwierząt laboratoryjnych.
Wyszukiwarka
Podobne podstrony:
zachowanie się zwierząt, biologia, Biologia I rok, Zachowanie się zwierząt
Biologiczne mechanizmy zachowania się zwierząt wy 1
Znaczenie percepcji wzrokowej w zachowaniu się zwierząt
Pytania na egzamin z BMZ, Studia UG, Psychologia, Semestr 1, Biologiczne mechanizmy zachowania się l
Bestia zachowuje się źle rozdział 4
PIERWSZA POMOC, Zachowanie się podczas wypadku samochodowego 2
wierszyki Dobre wychowanie[1], RÓŻNE, ZASADY ZACHOWANIA SIĘ W SZKOLE
Skuteczne zachowanie się w sytuacjach konfliktowych, psychologia - nowe
Zachowanie się substancji szkodliwych w glebie
Prawo administracyjne (11 stron), Prawo administracyjne - zbiór norm regulujących organizacje i zach
Bestia zachowuje się źle rozdział 3
Bestia zachowuje się źle Rozdział 7
3 6 Umiejętność zachowania się w sposób poprawiający wizerunek przewoźnika
Bestia zachowuje się źle rozdział 14
Bestia zachowuje się źle Rozdział 10
więcej podobnych podstron