Podział dziejów Ziemi:
ERY OKRESY CZAS TRWANIA
Kenozoiczna Czwartorzęd, trzeciorzęd 65
Mezozoiczna kreda, jura, tria 165
Paleozoicz perm, karbon, dewon, sylur,
ordowik, kambr 370
Proterozoiczna ok. 2000
Archaiczna ok. 2400
PRZYPUSZCZAMY, CO BYŁO POTEM
Nie jest wykluczone, że prób opanowania lądu było wiele. Wśród sylurskich skamielin znajdują się organizmy łączące w sobie cechy roślin lądowych i glonów, jednak nie mające nic wspólnego z innymi, znanymi nam roślinami lądowymi.
Do niedawna wszystkie prymitywne rośliny lądowe, których szczątki znajdowano w osadach pochodzących z przełomu syluru i dewonu, zaliczano do jednej grupy psylofitów. W miarę doskonalenia technik badawczych okazało się, że są one niezwykle zróżnicowane. Wkrótce stwierdzono, że tzw. psylofity to właściwie kilka zupełnie odmiennych grup roślin, 'na siłę' połączonych w jedną. Zarzucono więc nazwę psylofity, choć można ją spotkać jeszcze w starszych podziałach systematycznych i w literaturze. Miejsce psylofitów zajęły trzy różne, lecz blisko spokrewnione ze sobą grupy roślin: ryniofity, zosterofilofity i trymerofity.
Ryniofity - początek ery roślin lądowych
Ryniofity to najpierwotniejsze, pionierskie rośliny lądowe. Pojawiły się na ziemi w pierwszej połowie ery paleozoicznej - w sylurze. Zapoczątkowały wszystkie linie rozwojowe prowadzące do współcześnie żyjących lądowych gatunków roślin.

Do ryniofitów należą, uważane za pierwsze, najprostsze i najbardziej prymitywne, lecz typowo lądowe rośliny - kuksonie, żyjące pod koniec syluru. Ich wzniesione nagie, zakończone pojedynczymi zarodniami, widlasto rozgałęziające się pędy miały kilkanaście centymetrów wysokości. Miały one zaczątki prymitywnych wiązek przewodzących. Najlepiej jednak znane i zbadane są ryniofity odkryte w skamieniałych dewońskich bagniskach odkopanych pod szkocką miejscowością Rhynie (stąd nazwa całej grupy). 
Należą do nich rynia i hornea.
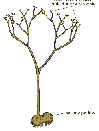
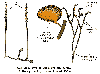
Dorastały one do pół metra wysokości. Ich rozgałęziające się widlasto pędy wyrastały z płożącego się kłącza o podobnej budowie. Nie wykształciły korzeni. Ich łodygi były zakończone zarodniami - były to więc sporofity. Równie prosta była budowa wewnętrzna tych roślin; ciało ich było pokryte epidermą, pod którą znajdowała się tkanka miękiszowa pełniąca funkcje asymilacyjne. Środkiem przebiegała cienka wiązka przewodząca. W tych samych bagniskach odkryto też rośliny będące gametofitami, mające zbliżoną do rynii budowę zewnętrzną i wewnętrzną. W cyklu życiowym ryniofitów najprawdopodobniej zarówno sporotit, jak i gametofit były pokoleniami równoznacznymi.
{kind=link}
Ewolucja roślin lądowych przebiegała bardzo szybko. W ciągu 40 mln lat (to się nazywa szybko...) z ryniofitów utworzyły się wszystkie linie rozwojowe współczesnych roślin lądowych. Dały one początek z jednej strony grupom roślin o dominującym gametoficie - mszakom i glewikom, zaś z drugiej roślinom, u których w cyklu życiowym dominuje sporofit, zaś gametofit uległ niejednokrotnie znacznej redukcji i całkowitemu uzależnieniu od pokolenia bezpłciowego. Ta gałąź rozwojowa doprowadziła do powstania współczesnych widłakowych, skrzypów, paproci i roślin nasiennych. Same ryniofity wymarły pod koniec dewonu, 370 mln lat temu.
Zosterofilofity - praprzodkowie współczesnych widłaków
Wywodzące się z ryniofitów zosterofilofity dały początek linii rozwojowej prowadzącej do współczesnych widłakowych.
{kind=link}
Zosterofilofity pojawiły się na Ziemi na początku dewonu. Ich przodkami były ryniofity. Odróżniały się znacznie od pozostałych grup roślin. Ich nerkowate zarodnie osadzone na trzoneczkach zebrane były na końcach bezlistnych pędów.
Zosterofilofity były niewielkimi roślinami tworzącymi gęste darnie. Ich gametofity i sporofity odgrywały w cyklu życiowym taką samą rolę. Zosterofilofity stanowią ogniwo łączące ryniofity z widłakami.
Trymerofity są pniem drzewa rodowego obecnych skrzypów, paproci i roślin nasiennych
Wywodzące się z ryniofitów trymerofity zapoczątkowały linię rozwojową skrzypów, paproci i pranagozalążkowych - bezpośrednich przodków roślin nagonasiennych.
Szczątki trymerofitów znamy z osadów dewońskich. Były bezlistnymi roślinami, pochodzącymi bezpośrednio od ryniofitów. U prymitywnych trymerofitów łodygi rozgałęziały się widlasto. Zwykle jednak jedno z rozgałęzień rosło wolniej, było mniejsze i 'spychane' na bok przez drugie, rosnące mocniej i szybciej. W ten sposób doszło do wykształcenia wyraźnej osi głównej. Tak ukształtowana roślina była mechanicznie silniejsza i odporniejsza na złamania od widlasto rozgałęzionych ryniofitów i części zosterofilofitów. Zarodnie wykształcone na końcach słabiej rosnących gałązek nie hamowały wzrostu pędu głównego.
Trymerofity mogły stać się gigantami ówczesnej flory - dorastały do l m wysokości. Gałązki boczne stawały się coraz krótsze, zachodziły na siebie, aż w końcu skupiały się w pęczki. Jednocześnie następowała ich specjalizacja. U następców trymerofitów na końcach jednych z nich wykształcały się zarodnie - były to liście zarodnionośne, czyli sporofile. Inne odgałęzienia zbliżały się do siebie i zrastały, przybierając postać szerokich liści asymilacyjnych.
{kind=link}
Trymerofity zapoczątkowały dwa szeregi rozwojowe roślin lądowych: skrzypów i paproci oraz pranagozalążkowych, które dały początek przeważającej części żyjących współcześnie roślin lądowych.
ŻYCIE TOCZY SIĘ DALEJ...

Tempo i kierunki przebiegu ewolucji roślin są nierozerwalnie związane ze zmianami geologicznymi i klimatycznymi ówczesnej Ziemi. Początkowo, nieprzychylny wegetacji, suchy, pustynny klimat końca syluru (415 min lat temu) wymuszał na prymitywnych roślinach pozostawanie na terenach stale zalewanych wodą. Jednocześnie 'promował' wszelkie sposoby unikania skutków suszy. Efektem tych zmagań było pojawienie się w dewonie (400 min lat temu) i dalszy bujny rozwój roślin naczyniowych. Jednak ich tkanka przewodząca nie była jeszcze wystarczająco sprawna, nie mogły więc osiągać zbyt dużych rozmiarów. Nie mogły także wykształcić liści o dużych powierzchniach, narażonych na zbyt silne parowanie. Ich rozmnażanie się było ciągle uzależnione od wody, dlatego u wszystkich żyjących wówczas, dominujących grup roślin zaznaczyła się tendencja do ochrony gametofitów i młodych sporofitów w specjalnych osłonach.
Nie bez znaczenia było także osiągnięcie stanu, w którym młody sporofit mógł przerwać na pewien czas swój rozwój i pozostać w osłonach, co dawało mu szansę przetrwania ewentualnych niekorzystnych zmian w środowisku. Doprowadziło to do powstania prymitywnych roślin nasiennych. Po gwałtownych ruchach tektonicznych w dewonie (nie poradziły sobie z nimi pierwotne ryniofity, zosterofilofity i trymerofity) nadszedł czas wilgotnego i bagiennego karbonu (345 min lat temu). W tym właśnie czasie na lądach panowali potomkowie wymarłych grup: wielkie widłakowe i skrzypowe, a także nieco mniej okazałe paprociowe. Rozwijały się w dalszym ciągu rośliny nasienne. Wielkie i gwałtowne zmiany klimatyczne i geologiczne w permie (270 min lat temu) spowodowały wyginięcie olbrzymiej części paleozoicznych gatunków. Powstawały nowe łańcuchy górskie, klimat uległ drastycznemu ochłodzeniu. Z Antarktydy zeszły lodowce, które pokryły większą część półkuli południowej. Wskutek obniżenia się temperatury wody wymarło też wiele gatunków glonów i zwierząt morskich (dziwne, że po czymś takim życie w ogóle nie musiało powstawać od nowa...). Pozostały te organizmy, które najlepiej poradziły sobie z przeciwnościami natury. W nowo rozpoczynającej się erze mezozoicznej przewagę osiągnęły, pozbawione konkurencji wielkich paprotników, rośliny nagonasienne.
Mszaki są grupą roślin, w których cyklu życiowym dominuje gametofit
Do mszaków zaliczane są rośliny o różnorodnej budowie, których wspólną cechą jest wykształcanie zielonego, samodzielnego gametofitu, dominującego nad uzależnionym od niego, często bezzieleniowym sporofitem. Nie wykształcają typowej tkanki przewodzącej.
Gametofit mszaków jest zieloną rośliną o zróżnicowanej, charakterystycznej dla poszczególnych grup, niejednokrotnie złożonej budowie. Może być on plechowaty bądź składać się z łodyżki, liści i chwytników. W budowie anatomicznej możemy wyróżnić tkanki przewodzące (są one bardzo prymitywne, zbudowane inaczej niż u roślin naczyniowych, zaś ich efektywność jest znikoma), tkankę miękiszową i okrywającą. Stopień ich wykształcenia i budowa są bardzo różnorodne u poszczególnych grup mszaków. Gametofit zaopatruje uzależniony od niego sporofit w pokarm, wodę i sole mineralne.
Sporofit mszaków zwykle jest pozbawiony możliwości fotosyntezy i całkowicie uzależniony od gametofitu; żyje od niego krócej. Zbudowany jest z łączącej go z gametofitem stopy i prostej, nie rozgałęzionej 'łodyżki', zwanej setą, zakończonej pojedynczą zarodnią. Wnętrze zarodni wysłane jest warstwą komórek zdolnych do podziałów mejotycznych. Z każdej z nich, w wyniku mejozy, powstają cztery haploidalne zarodniki. Po dojrzeniu i wysypaniu się spór z zarodni sporofit ginie. 
Przemiana pokoleń przebiega we wszystkich grupach mszaków według jednego schematu.
Na powierzchni gametofitu rozwijają się gametangia żeńskie - rodnie i gametangia męskie - plemnie. W wyniku podziałów mitotycznych w rodniach powstają haploidalne gamety żeńskie - komórki jajowe, zaś w plemniach gamety męskie - ruchliwe, uwicione plemniki. Aby mogło nastąpić zapłodnienie, plemnik z plemni musi przedostać się do rodni. Może tego dokonać jedynie w środowisku wodnym - może być to kropla deszczu lub warstwa rosy, jednak bez wody zapłodnienie jest niemożliwe.
Wynikiem zapłodnienia jest diploidalna zygota, od której rozpoczyna się rozwój sporofitu.
Zarodniki niektórych gatunków mszaków mogą być identyczne pod względem morfologicznym, lecz mają zdeterminowaną pleć. Z jednych rozwijają się gametofity wyłącznie męskie z plemniami, z drugich - żeńskie z rodniami. U innych gatunków zarodniki są identyczne także pod względem fizjologicznym - rozwijają się z nich gametofity obupłciowe, mające zarówno plemnie, jak i rodnie.
Gdy zarodniki padną na odpowiednie podłoże, kiełkują w bryłowaty lub nitkowaty zaczątek gametofitu, rozwijający się w haploidalny gametofit.
Mszaki mogą rozmnażać się bezpłciowo także przez rozmnóżki - kilkukomórkowe utwory o zgrubiałych ścianach.
GAMETOFITY MCHÓW MAJĄ POSTAĆ ULISTNIONEJ ŁODYŻKI
Gametofit mchów składa się z ulistnionej łodyżki i chwytników. Organy te pełnią podobne funkcje jak łodygi, liście i korzenie roślin naczyniowych. Powstają jednak w inny sposób, są też odmiennie zbudowane. Wnętrze łodyżek mchów tworzą komórki miękiszu. Niekiedy pojawiają się specyficzne tkanki przewodzące. Liście mchów właściwych zbudowane są z jednej warstwy komórek. Wielokomórkowe chwytniki przytwierdzają roślinę do ziemi. Zaopatrują ją w wodę i sole mineralne.
Tkanki przewodzące mchów zbudowane są głównie z martwych, wydłużonych komórek - hydroidów, pełniących takie funkcje jak naczynia w drewnie roślin naczyniowych. Czasem wokół wiązki hydroidów biegnie tkanka przewodząca zbudowana z żywych komórek - leptoidów - o budowie przypominającej bardzo prymitywne łyko.

Sporofit mchów jest organizmem stosunkowo trwałym (u niektórych gatunków może żyć nawet do 3 lat); zawsze jednak jest uzależniony od gametofitu i ściśle z nim związany. Początkowo sporofit rozwija się w rodni. W miarę wzrostu przerywa on jej ścianki i rosnąc podnosi na sobie jej górną część jako tzw. czapeczkę, ochraniającą młodą zarodnię. Dojrzały sporofit składa się ze stopy łączącej go z gametofitem i sety, na której wyniesiona jest zarodnia. W środku zarodni znajduje się, charakterystyczna dla mchów, kolumienka. Zarodnia zamknięta jest wieczkiem. Odpada ono po dojrzeniu zarodników, otwierając zarodnię i umożliwiając ich wysyp.
Zarodnik kiełkuje w nitkowaty bądź plechowaty zaczątek gametofitu, zwany splątkiem.
Po odpadnięciu wieczka brzeg powstałego otworu otaczają zębiaste wyrostki, tzw. ozębnia. Gdy jest mokro, składają się one do środka zarodni uniemożliwiając wysyp zarodników, które w wilgotnym powietrzu zlepiałyby się i nie mogły oddalić od macierzystych roślin.
|
CO WARTO ZAPAMIĘTAĆ |
Gametofit mchów ma postać ulistnionej łodyżki z chwytnikami. U niektórych mchów gametofit może mieć zróżnicowaną na tkanki budowę anatomiczną. Dojrzały sporofit składa się ze stopy, sety i zamkniętej wieczkiem zarodni. Może rozwijać się i żyć nawet kilka lat. Zarodnik kiełkuje w splątek, który jest zaczątkiem gametofitu. |
Prątniki (mchy właściwe) są mchami najczęściej spotykanymi wśród roślin dna lasu

Gametofit prątników składa się z ulistnionej łodyżki i chwytników. Wzdłuż, zbudowanych z jednej warstwy komórek, blaszek liści biegnie zwykle pasmo kilku warstw komórek, zwane żebrem lub nerwem.
Stanowi ono tkankę przewodzącą liścia, nie łączącą się jednak z wiązką przewodzącą łodygi. Sporofit ma dobrze wykształconą setę, zaopatrzoną w wiązki przewodzące, rozwijającą się na długo przed wykształceniem się zarodni. Zarodnia, wyniesiona wysoko nad gametofit, ma stosunkowo złożoną budowę wewnętrzną. Otwiera się wieczkiem.
Z zarodnika kiełkuje nitkowaty, wielokomórkowy splątek. Daje on początek wielu ulistnionym łodyżkom.
Prątniki i lasy

Prątniki występują w wielu różnorodnych siedliskach. Można je spotkać na nagich skałach, korze drzew (w krajach tropikalnych nawet na liściach), porastają betonowe chodniki i eternitowe pokrycia dachów. Są jednymi z pionierskich roślin zasiedlających, wykorzystujących i przekształcających nieprzyjazne dla roślin wyższych środowiska. Rosną także w wodzie, zarówno bieżącej, jak i stojącej. Nie ma ich jedynie w morzach, a na terenach zasolonych występują niezwykle rzadko.
Najczęściej jednak mchy właściwe spotkać można w najniższym piętrze lasów iglastych i (w mniejszym stopniu) liściastych, gdzie budują tzw. warstwę mszystą. 

Rola mchów w życiu lasu jest ogromna - magazynują one w swych darniach wodę, przyczyniając się do powstania specyficznego leśnego mikroklimatu, niezbędnego do funkcjonowania tych olbrzymich, niezwykle złożonych formacji roślinnych. Zatrzymując wodę leśne mchy sprawiają, że nawet po długotrwałych, intensywnych opadach nie spływa ona natychmiast do rzek, lecz przesiąka do nich stopniowo - poziom wód nie podnosi się wówczas tak szybko, niknie groźba gwałtownych powodzi. Warstwa mchów i butwiejącej ściółki stanowi też środowisko życia olbrzymiej liczby pierwotniaków, grzybów oraz zwierząt bezkręgowych (np. wrotków, nicieni, różnych stawonogów czy ślimaków) i drobnych kręgowców (np. żab, jaszczurek, gryzoni).

Dla wielu z tych organizmów stanowi ona jedyne siedlisko, jakie znają - tu rodzą się, dojrzewają, wydają potomstwo i giną. Między innymi dlatego tak ważne jest, aby w lesie nie niszczyć bezmyślnie ściółki i mchów.
|
CO WARTO ZAPAMIĘTAĆ |
Gametofit mchów właściwych zróżnicowany jest na łodyżkę, liście i chwytniki. W jego budowie anatomicznej można wyróżnić prymitywne tkanki przewodzące. Blaszki liści prątników mają żebro (nerw środkowy) - charakterystyczne zgrubienie powstałe z tkanki przewodzącej. Mchy właściwe są głównym składnikiem warstwy mszystej dna lasów i jednym z elementów warunkujących równowagę ekologiczną tych zbiorowisk. |
Torfowce są bardzo wyspecjalizowanymi mchami, dzięki którym istnieją osobliwe formacje roślinne - torfowiska

Torfowce są mchami o szczególnej budowie gametofitu. Łodyżka torfowca ma zdolność nieograniczonego wzrostu, przy czym rośnie tylko jej część wierzchołkowa, dolna zaś stopniowo obumiera i butwieje. Liście wyrastają na gałązkach bocznych i bezpośrednio na łodydze. Zbudowane są z jednej warstwy komórek, wśród których możemy wyróżnić zielone komórki asymilacyjne i tzw. komórki wodonośne.

Komórki te są duże, martwe, a ich ściany są silnie zgrubiałe i porowate. Podobne komórki wykształcają się też na łodyżce. Dojrzałe torfowce nie mają wiązek przewodzących ani chwytników.

Rodnie i plemnie torfowców wyrastają w szczytowej części gametofitu na bocznych gałązkach, dzięki temu nie jest hamowany ciągły wzrost głównej łodyżki.
Sporofit torfowców ma bardzo krótką setę. Na jej szczycie znajduje się duża, kulista zarodnia otwierająca się wieczkiem.

W zarodni torfowców nie ma ozębni. Po dojrzeniu zarodników kolumienka pęcznieje. Dzięki temu w zarodni zaczyna wzrastać ciśnienie, pod wpływem którego odrywa się wieczko i zarodniki zostają wyrzucone. Kiełkują one w plechowate splątki. Splątek torfowców może rozmnażać się wegetatywnie, wytwarzając nitkowate wyrostki, z których rozwijają się nowe, plechowate splątki. Młode gametofity przytwierdzają się do podłoża chwytnikami, które po pewnym czasie zanikają.
Torfowce, torfowiska, człowiek i historia
Dzięki dużej ilości komórek wodonośnych torfowce posiadają olbrzymią zdolność chłonięcia wody - mogą jej zmagazynować nawet 25 razy tyle, ile same ważą. W naturze rosną zawsze gromadnie, tworząc zwarte, nasycone wodą płaty. Można je spotkać w wilgotnych borach sosnowych i świerkowych.

Największą rolę w przyrodzie i gospodarce człowieka odgrywają torfowce budujące niezwykłe zbiorowiska roślinne - torfowiska.
W silnie wilgotnych miejscach, gdzie stale gromadzi się woda, np. w dolinach powoli płynących, meandrujących rzek, pozbawionych odpływu zagłębieniach terenu, przy płytkich brzegach leśnych jezior o czystej wodzie torfowce znajdują znakomite warunki rozwoju. Rosną szybko (przybywa im około 6 cm długości rocznie) i równie szybko się rozmnażają. Pokrywają one swe siedliska grubym, stale narastającym kożuchem nasączonym wodą. Jednocześnie dolne części roślin obumierają i w wysyconym wodą, pozbawionym tlenu środowisku, przy udziale grzybów i bakterii, ulegają powolnemu rozkładowi. Powstaje z nich torf - masa w różnym stopniu rozłożonych części roślin, wysycona szczególnymi związkami organicznymi i mineralnymi (tzw. humusem), o kwaśnym odczynie, wiążąca duże ilości wody.
Torfowiska powstają na obszarach o klimacie charakteryzującym się dużą przewagą opadów nad parowaniem i stosunkowo niską temperaturą, hamującą parowanie i zbyt szybki rozkład materii organicznej (przy szybkim rozkładzie i braku wody, w obecności tlenu, szczątki roślinne mogłyby ulec całkowitej przemianie na rozpuszczalne w wodzie związki mineralne; nie powstałyby wtedy złoża torfu). Torfowiska najczęściej spotykane są w strefie tajgi i tundry.
Stanowią jeden z głównych elementów tworzących krajobrazy krain Północy, nadając im niezwykły, surowy koloryt. W Polsce są zbiorowiskami bardzo rzadkimi, zasługującymi na troskliwą ochronę. Najwięcej znajduje się ich na Pomorzu, Suwalszczyźnie, Podlasiu i Polesiu, czyli na północy i wschodzie kraju.

Torfowiska są ostojami wielu gatunków roślin niespotykanych w żadnych innych zbiorowiskach. Najpospolitszymi z nich są m.in.: będące pod ochroną bagno zwyczajne (używane do zwalczania moli) - i żurawina błotna (wyrabia się z niej znakomitą konfiturę dodawaną do pieczonych mięs).
Wiele roślin jest przedstawicielami gatunków w naszym kraju zagrożonych, rzadkich, ginących i chronionych. Należą do nich m.in. owadożerne rosiczki, wiele gatunków turzyc i storczyków,. Rozległe i niedostępne tereny bagienne są ostoją wielu chronionych, zagrożonych i ginących gatunków zwierząt, np. ptaków (m.in. żurawi, czarnych bocianów, batalionów, niezwykle rzadkich wodniczek i derkaczy, drapieżnych sów błotnych, orlików i błotniaków) i ssaków (m.in. łosi, wydr, bobrów, wilków) oraz całej rzeszy zwierząt bezkręgowych.
Stosunkowo żyzne torfowiska, leżące w dolinach rzek i zasilane ich wodami (tzw. torfowiska niskie), po odpowiednio przeprowadzonym odwodnieniu są wykorzystywane rolniczo jako łąki i pastwiska. Zbytnie osuszenie torfowisk powoduje jednak obniżenie poziomu wód gruntowych na terenach zarówno z nimi sąsiadujących, jak i znacznie odległych, zwłaszcza gdy gleby wokół są łatwo przepuszczalne i piaszczyste. Prowadzi to do zmiany warunków klimatycznych całego regionu. Niejednokrotnie źle przeprowadzona melioracja torfowisk spowodowała więcej długotrwałych i nieodwracalnych szkód, niż dała doraźnych korzyści.
Torf wykorzystywany jest ponadto w ogrodnictwie jako dodatek spulchniający ziemię, w lecznictwie -jako tzw. borowina oraz do wyrobu leków; wyciągi z torfu są także podstawowym składnikiem niektórych kosmetyków.
Torf ma właściwości konserwujące. Wiele znalezisk archeologicznych miało miejsce na terenach dawnych lub jeszcze istniejących torfowisk, gdzie w bardzo specyficznych warunkach, przy braku tlenu, w obecności kwasów organicznych, zachowały się niemal nie naruszone ciała zwierząt i ludzi z dawnych epok. Pokłady torfu kryły m.in. gród w Biskupinie (Kujawy) - niezwykły archeologiczny wizerunek codziennego życia ludzi sprzed ponad 2,5 tysiąca lat.
|
GAMETOFITY WĄTROBOWCÓW SĄ PLECHOWATE BĄDŹ MAJĄ POSTAĆ ULISTNIONEJ ŁODYŻKI

Gametofit wątrobowców może mieć postać plechy albo ulistnionej łodyżki. Przytwierdzony jest do podłoża jednokomórkowymi chwytnikami. Plechy mogą przybierać różne kształty. Łodyżki mogą pokładać się lub wznosić, być wielokrotnie rozgałęzione lub proste, dwustronnie spłaszczone. Listki są zawsze jednowarstwowe, bez nerwu środkowego (tak charakterystycznego dla mchów) i są zawsze ułożone płasko wzdłuż łodyżki. Niekiedy po stronie spodniej łodygi występuje rząd listków przekształconych w kubkowate twory służące do magazynowania wody.
Budowa wewnętrzna plech i ulistnionych łodyżek może być bardzo prosta bądź zróżnicowana na tkanki: okrywającą, spichrzową, wzmacniającą, asymilacyjną i przewodzącą. Komórki większości wątrobowców zawierają specyficzne twory, zwane ciałkami oleistymi. Ich rola nie jest dotychczas wyjaśniona.
Sporofit wątrobowców jest organizmem bardzo delikatnym i nietrwałym. Pozostaje on w rozrośniętej, spodniej części rodni, z której zaczyna wyrastać tuż przed wysypem zarodników. Zarodnia otwiera się przez podłużne pęknięcia ścian lub rozpada na nieregularne fragmenty, rzadko jest opatrzona wieczkiem. Wnętrze zarodni jest wypełnione zarodnikami i charakterystycznymi dla wątrobowców komórkami pozbawionymi protoplastu, zwanymi sprężycami lub elaterami. W zależności od wilgotności powietrza skręcają się one lub rozkręcają, ułatwiając wysiew zarodników.
Zarodnik kiełkuje w bryłowatą masę komórek, która rozwija się w pojedynczy gametofit. U wątrobowców brak typowo wykształconego splątka.
Wątrobowce w przyrodzie
W naturze wątrobowce nie są zbyt rozpowszechnione, występują jednak na całej kuli ziemskiej; od obszarów arktycznych po wilgotne lasy tropikalne, gdzie jest ich największa różnorodność. Podobnie jak mchy, mogą zasiedlać zarówno miejsca bardzo suche, jak i występować w zbiornikach wodnych. Spotykane są na korze drzew, a w krajach tropikalnych także na liściach. Wiele z nich należy do grupy roślin pionierskich.
Jeden z naszych najpospolitszych krajowych gatunków, plechowatą porostnicę wielokształtną (Marchantia polymorpha), często można spotkać na wilgotnej glebie, stale zwilżanych skałach, a także w miejscach po świeżo wypalonych ogniskach. Początkowo jest tam tak duże stężenie soli mineralnych, że większość roślin nie jest w stanie się osiedlić. Wykorzystują to porostnice, którym zasolenie zbytnio nie przeszkadza, a jednocześnie nie muszą przez pewien czas obawiać się konkurencji. Plechowate wątrobowce żyją także w słodkiej wodzie. Ricciocarpus natans ma drobne, wielokrotnie widlasto porozgałęziane plechy, tworzące małe rozetki ścielące się po dnie. Podobnie wygląda wgłębka Riccia fluitans, pływająca po powierzchni wody, tworząca na niej gęste, gąbczaste kożuchy. Jest to dobrze znany akwarystom pospolity 'mech wodny', którego skupienia są ulubionym schronieniem narybku i drobnych rybek.
Większość wątrobowców tworzących plechy należy do rzędu Marchantiales (porostnicowatych), zaś tworzących łodyżki - do rzędu Jungermaniales, gromadzącego większość gatunków wątrobowców.
Wyszukiwarka
Podobne podstrony:
Ćwiczenie 1, Ćwiczenie 1 (3), Chronologia dziejów ziemi :
Scenariusz lekcji z dziejów ziemi wyrzyskiej?resowany jest przede wszystkim do nauczycieli z m2
Karta pracy klasa V podział dziejów ludzkości, pedagogika, dydaktyka, scenariusze, konspekty, karty
Dzieje człowieka na przestrzeni dziejów Ziemi, Matura na 100%
ciężkowski,hydrogeologia, Chronologia dziejów Ziemi
ciężkowski,hydrogeologia, OGÓLNY PODZIAŁ I HISTORIA ZIEMI
Dzieje ziemi- podziałna ery i okresy, NAUKA, geografia, Geografia(1)
Geologia (rok II), GEOLOGIA, Podział geologii: dyscyplina wiedzy zajmująca się nauką o Ziemi
Bogaczewicz,hydrologia i nauka o ziemi, podział cieków
Pułk Piechoty Ziemi Ziemi Wadowickiej (Z dziejów tradycji i nazwy 12 pułku piechoty)
Definicja i podzia skazy krwotocznej
Podział skał
KLIMATY ZIEMI
Określenie terminu ekologia Podział ekologii z uwzględnieniem
02 Pojęcie i podziały prawaid 3482 ppt
podziały złamań cz2 1sd
więcej podobnych podstron