DSCN6077 (Kopiowanie)

*V\
%■
jak zauważył to już Hneckcl - jamochłony sq dwuwarstwowe (zbudowane z dwóch waislw komórek z zewnętrznej cktodermy i wewnętrznej entodermy). Łotwo zatem możemy sobie wyobrazić ich powstanie t pojedynczej warstwy komórek pierwotnego organizmu kolonialnego, jak to ilustruje ryc. 2-10 i jak to możemy obserwować w rozwoju embrionalnym większości bezkręgowców i niektórych kręgowców. Natomiast stojące wyżej od nich ewolucyjnie zwierzęta snlrójwarstwowe. tzn. pomiędzy ektodermą i entodermą pojawia się trzeci listek -mczodenna. Nie znaleziono do tej pory żadnych skamieniałości organizmów pośrednich, co można zrozumieć, gdyż zwierzęta te nie wytwarzają żadnych szkieletów, a ich miękkie ciała nie pozostawiają śladów. Płazińce posiadają symetrie dwuboczna. jamochłony zaś
•*S|
ydćiowco
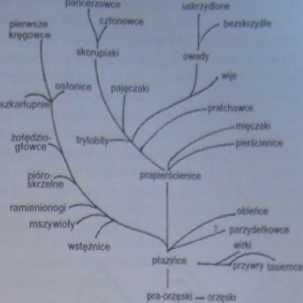
Ryc. 2-37. Pokrewieństwa ewolucyjne bezkręgowców (JD)
(2*ł °***

____ _ ')■ Sądzi się jednak, że symetria
promienista jest przystosowaniem do osiadłego życia w morzach - gdy zarówno pokarm, jak i zagrażające niebezpieczeństwo może się pojawić z każdej strony.
Obleńce (typ Aschelminlhes) w porównaniu z płazińcami, od których pochodzą, osiągnęły dwie korzystne cechy. Po pierwsze - wykształciły jamę ciała wypełnioną płynem, co umożliwia większą swobodę poruszania się. Po drugie - ich przewód pokarmowy ma kształt rury i posiada dwa otwory: gębowy i odbytowy, co znacznie polepsza wydajność procesów wchłaniania i trawienia pokarmów.
Pierwotne płazińce dały prawdopodobnie początek jeszcze dwu grupom zwierząt - pierwotnym pierścienicom i wsteżnicom.
Pierwotne pierścienice - które być może podobne były do współczesnych - uważa się za przodków trzech typów zwierząt: pierścienic. (Annelida), mięczaków (Molhisca) i stawonogów (Artliropóda). Część badaczy uważa jednak, że zarówno pierścienice, jak i wstężnice (Nemertim) mogły mieć wspólnego nieznanego przodka. Cechą wspólnądla pierścienic, wielu mięczaków, a także wężnic, szkartupni (typEchinodermata) i mszywiołów (Baryozoa) jesl wytwarzanie w toku rozwoju utzęsionej, pływającej larwy zwanej trochoforą. Trochofory wszystkich wymienionych typów mają podobną budowę (ryc. 2-38).
Choć mięczaki - w odróżnieniu od pierścienic - nie mają segmentowanego ciała, to jednak jednym z dowodów na wspólne pochodzenie tych dwóch typów jest żyjący obecnie reliktowy mięczakNeopilina (wydobyty w 1952 roku z dna Pacyfiku). Zwierzę to którego najbliżsi krewni wymarli w ordowiku - wykazuje segmentację ciała i zapewne stanowi ogniwo pośrednie między pierwotnymi pierścienicami a mięczakami. Pierwsze skamieniałości mięczaków znamy z kambrn. Jak panuętamy, typ ten był liczebny we wszystkich epokach geologicznych, a i dzisiaj jest jednym i trzech dominujących na Ziemi (100 000 gatunków).
Pochodzenie stawonogów od pierścienic zazwyczaj nie jest kwestionow ane (ryc. 2-37). Już tu pww wy rzut oka widać wspólną dla obu typów, dwuboczną symetrię segmentów anego ciała Najliczniejszymi skamieniałościami stawonogów z okresu luunbru satrylobitylryc. 2-12). Ich tody nawiązują zarówno do pierścienic, jak i do skorupiaków i pajęczaków. Jedynym morskim
Wyszukiwarka
Podobne podstrony:
DSCN6077 (Kopiowanie) teąitf (2n)‘*aZ lik rauwtóyl to już Hncckcl jamochłony są dSimiStoUKS (zbudowa
Zauważali to już starożytni filozofowie (np. Arystoteles), definiując człowieka jako „istotę
8 9 (5) To wszystko, co odnosiło się do „naukowości” w sensie ogólnym, jak na to już wskazywaliśmy,
(zanim państwo coś zauważy, to już się to dzieje). Ogólnie każde państwo stosuje interwencjonizm, je
DSC?23 (2) (jak zauważył to Bazin) nie jest kadrowaniem, ale przysłonięciem, osoba, która z niego wy
skanuj0038 (116) Rozdział 4 mNAWIĄZYWANIE KONTAKTU Z DUCHEM OPIEKUŃCZYM Jak to już zostało wcześniej
20130 Zdjęcie0238 (3) ■tu Huzdoal
PRZEGLĄD PIŚMIENNICTWA. 277 skiej, jak tylko uczestniczy! w ułożeniu i podpisaniu tego adresu, to ju
Drzewo życia 7 szybciej i łatwiej to spostrzegamy, jeśli — jak zauważają autorzy cytatu — istotnie o
mię. jak zauważył Patek, nie pozostanie to bez wpływu na sytuację na Zachodzie. Podkreślono, ze „Man
DSC05712 (3) LII Piotr Baryktt Co by mię każdy pomniał; wiecie jak? — Piwowym. 530 To już mam to wsz
więcej podobnych podstron