DSCN6381 (Kopiowanie)

f. Przegląd świata biot tywych I Ich klasyfikacja 77
7.4.5.I. Główne tendencje ewolucyjne u roślin
Jeden z głównych kierunków ewolucyjnych w święcie roślinnym to przejście od gatunków, u których w przemianie pokoleń przeważa wielkością gametofit do tych, u których przeważa sporofit (zaś gametofit zredukowany jest niejednokrotnie do kilku-kilkunastu komórek). O tej tendencji była już mowa w rozdz.2; przypomnijmy sobie ryciną 2-35. Możemy domyślać się, dlaczego owa tendencja ma miejsce. W komórkach haploidalncgo gametofitu występuje pojedyncza liczba genów (n). A zatem, gdy któryś z owych genów ulegnie niekorzystnej mutacji lub gdy zaistnieją niesprzyjające dlań zmiany środowiska, komórka (względnie cala roślina) musi zginąć. W organizmie diploidalnym każdy gen ma swój allcliczny odpowiednik, więc w przypadku wypadnięcia funkcji genu jego rolę może przejąć gen komplementarny. Im dłużej żyje dany osobnik, tym większe znaczenie ma dla niego możliwość uniknięcia niepożądanych mutacji - a zatem przewagę uzyskiwały te rośliny, u których pokolenie sporofitowe było bardziej długowieczne niż gametofitowe.
Druga ważna ewolucyjnie tendencja ma "na celu” doskonalenie zaopatrywania wszystkich komórek w wodę i sole mineralne. Większość mszaków nie posiada tkanek przewodzących, lecz u płonnika (Poliirychum) występuje w łodyżce prymitywna wiązka przewodząca. Poczynając od paprotników mamy już do czynienia z łykiem i drewnem, zaś rośliny okrytonasienne wykształciły naczynia (rozdz. 4.5.). Wiązki przewodzące pojawiły się najprawdopodobniej najpierw w sporofitach, a zatem osobniki tego pokolenia mogły osiągać większe rozmiary.
Ważny kierunek rozwojowy stanowi uniezależnienie rozrodu płciowego od obecności wody, co było jednym z warunków pełnego opanowania środowiska lądowego. Rośliny wodne oraz część roślin lądowych wytwarzają opatrzone wicią plemniki, które poruszają się w wodzie w kierunku komórki jajowej; takie rozwiązanie występuje jeszcze u sagowców i miłorzębowych, a więc roślin nasiennych. Jednak zasadniczy przełom dokonał się u roślin nasiennych, które wytworzyły pyłek. Ziarna pyłku są bardzo lekkie i mogą być na odległość setek kilometrów przenoszone przez wiatr, albo też (na mniejsze odległości) przez owady. Rośliny iednopienne mają na tym samym okazie zarówno męskie, jak i żeńskie narządy płciowe (np. sosna). U nich - podobnie jak i u roślin o kwiatach obupłciowych - możliwe jest samozapłodnienie. Stanowi ono rodzaj zabezpieczenia: jeżeli do komórki jajowej nie dotrze pyłek innych roślin, zygota może być zapłodniona własnym pyłkiem. Jednak samozapłodnienie - w przypadku, gdy powtarza się przez wiele pokoleń - może prowadzić do niekorzystnych zmian genetycznych, dlatego też wiele gatunków roślin wytworzyło mechanizmy przed nim zabezpieczające.
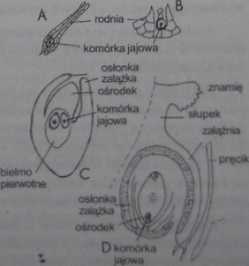
Wreszcie bardzo ważną tendencją ewolucyjną u roślin jest zabezpieczenie komórki iaiowei i zarodka przed bezpośrednim wpływem środowiska. U mszaków i paprotników komórka jajowa -choć unieruchomiona w rodni - ma bezpośredni kontakt z otaczającym środowiskiem, ponieważ rodnia nie jest zamknięta (ryc. 7-53). U nagonasiennych komórka jajowa oddzielona jest osłonką, zaś u okrytonasiennych aż dwiema warstwami; jedną stanowią warstwy zalążka, drugą - zewnętrzną - zalążnia. czyli zrośnięte podstawy słupka. Niedawno odkryto przedstawicieli nowej grupy roślin, u których komórka jajowa osłonięta jest aż trzema - o różnym pochodzeniu - warstwami.
Ryc. 7-53. Jeden z głównych kierunków ewolucyjnych u roślin to dążenie do zabezpieczenia komórki jajowej jak największą liczbą osłonek. U mszaków (A) i u paprotników, np. widliczki (B) komórka jajowa nie jest ukryła. U nagonasiennych (C) osłonięta jest przez zalążek: u okrytonasiennych (D) - przez Zalążek i zaiążnię <JD).
Wyszukiwarka
Podobne podstrony:
DSCN6381 (Kopiowanie) 1. Pnegląd Świnia istot żywych i ich klasyfikacja 77 7.4.5.I. Główne tendencje
DSCN6338 (Kopiowanie) 7. Przegląd świata biot żywych I ich klasyfikacja 7.3.1.1.1. Krótki przegląd g
DSCN6316 (Kopiowanie) 7.PRZEGLĄD ŚWIATA ISTOT ŻYWYCHI ICH klasyfikacja •4 gdy stworzyl Pan Bóg na Zi
DSCN6320 (Kopiowanie) E Przegląd świata istot żywych i ich klasyfikacja U Tabela 7-1. Klasyfikacja D
DSCN6326 (Kopiowanie) 7. Przegląd świata istot żywych i Ich klasyfikacja 17 Bardzo ważną cechą syste
DSCN6326 (Kopiowanie) 7. Przegląd świata istot żywych i ich klasyfikacje 17 Bardzo ważną cechą syste
DSCN6332 (Kopiowanie) Przegląd świata istot żywych i ich klasyfikacji 23 Pod ścianą komórkową znajdu
DSCN6334 (Kopiowanie) 7. Przegląd świata istot imirA i ich klasyfikacja 25 Różne gatunki bakterii ro
DSCN6336 (Kopiowanie) 7. Przegląd świata istot ijmyeh i ich klasyfikacja 27 Z medycznego punktu widz
DSCN6338 (Kopiowanie) 7. Przegląd świata Mol fywych I ich klasyfikacja 13.1.1.1. Krótki przegląd gro
DSCN6344 (Kopiowanie) ■ Pizegląd świata biot iynyrh i ich klasyfikacja 35 o haploidalncj liczbie ch
DSCN6346 (Kopiowanie) 7. Przegląd świata Istot żywych i ich klasyfikacja 37 Gr/yby zaliczano od czas
DSCN6350 (Kopiowanie) 7. Przegląd świata istot żywych I ich klasyfikacja 41 cytoplazmy jedno lub kil
DSCN6354 (Kopiowanie) 7. Przegląd świata istot żywych i ich klasyfikacja 45 7. Przegląd świata istot
DSCN6356 (Kopiowanie) 7. Przegląd świata istot żywych i ich klasyfikacja 47 szkól średnich i podstaw
DSCN6358 (Kopiowanie) 7. Przegląd świata istot żywych I ich klasyfikacja Ryc. 7-33. Piechy niektóryc
DSCN6360 (Kopiowanie) 7. Przegląd świata Istot żywych I Ich klasyfikacja 5J Tabela 7-X. Klasyfikacja
więcej podobnych podstron