DSCN6627 (Kopiowanie)

t Człowiek
t Człowiek
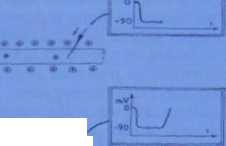
błony się wyrównują. Ten miejscowy stan depolaiyzacji określamy także jako potencjał generujący, ponieważ może on rozpocząć proces „reakcji łańcuchowej" - szerzenia się fali depolaryzacji wzdłuż aksonu. Jeżeli pobudzenie jest zbyt słabe (bodziec podprogowy). potencjał generujący nie osiąga wartości krytycznej, błona komórkowa usuwa jony sodu na zewnątrz i powraca stan polaryzacji (potencjał spoczynkowy). Jeżeli natomiast pobudzenie spowoduje osiągnięcie przez potencjał generujący pewnej wartości krytycznej, to stan depolaryzacji (otwarcie kanałów sodowych i wnikanie sodu do cytopłazmy) rozszerza się na błonę całej komórid; powstaje potencjał czynnościowy, przenoszony wzdłuż wypustek. Najmniejszy bodziec potrzebny do powstania potencjału czynnościowego nazywany jest bodźcem progowym. Potencjał czynnościowy przenoszony jest na zasadzie,, wszystko albo nic"—to znaczy albo powstaje i oznacza całkowitą depolaryzację błony neuronu, albo nie powstaje w ogóle.
Powyżej wartości progowej dalsze zwiększanie bodźca zawsze da jednaki efekt: depolaryzację, która zawsze jest całkowita i nie odzwierciedla siły czynnika pobudzającego.
Ryc. 8-61. Zmiany ładunków (stężenia jenów K' i Na') w nerwie i jego otoczeniu w procesie przewodzenia bodźca; a - polaryzacja (stan spoczynkowy); b - depolaryzacja (przebieg impulsu); c - polaryzacja ponowna (JD).
Do pojedynczej komórki nerwowej dochodzą nieraz setki wypustek innych neuronów, tworząc synapsy. Nawet jeżeli pojedyncze pobudzenie będzie niewystarczające dla powstania potencjału czynnościowego (bodziec podprogowy), może ono spowodować zmniejszenie potencjału spoczynkowego (część jonów sodu na moment wniknie w danym miejscu do komórki).
Takie zjawisko nazywamy torowaniem, gdyż ułatwia działanie następnych pobudzeń. Jeżeli w jednym czasie na komórkę zadziała wiele bodźców podprogowych. mogą się onezsumować i wywołać depolaryzację.
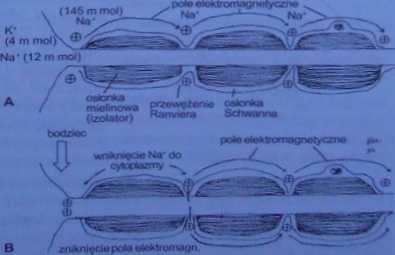
Ryc. 8-62. A - włókno zmielinizowane w sianie spoczynku: H - po zadziałaniu bodźca (opis w Ickścic)
Depolaryzacja szybko się rozprzestrzenia i trwa kilka milisekund. W tym czasie błona neuronu aktywnie usuwa na zewnątrz jony sodu, wymieniając je na wyrzucone jony potasowe. Czynność ta jest możliwa dzięki systemowi enzymów, określanych mianem pompy sodowo-potasowej. Do swego funkcjonowania wymaga ona energii pochodzącej z rozkładu ATP (uzyskiwanego w procesach metabolicznych komórki). Działanie pompy doprowadza do stanu repolaryzaćji. czyli przywrócenia potencjału spoczynkowego (ryc. 8-61 c). Oczywiście dopóki nie nastąpi ponowna polaryzacja błony, przewodzenie następnego impulsu jest niemożliwe; ten czas - nazywany absolutnej - wynosi od 0,4 do 2 milisekund.
Opisany powyżej sposób przenoszenia bodźca wzdłuż aksonu dotyczy włókien niezmielinizowanych, czyli nie posiadających osłonki rdzennej. Obecność osłonki mie-linowej, która pełni rolę izolatora — uniemożliwia polaryzację włókna. Jednak osłonka miclinowa nie jest ciągła, lecz składa się z pochewkowatych segmentów; każdy segment jest utworzony przez jedną komórkę glejową. Pomiędzy segmentami osłonki mielinowej znajdują się wąskie przerwy, zwane przewężeniami (wędami) Kanviera (ryc. 8-62). Odsłonięte części neuronu (np. perikarion, nagie zakończenia) oraz przewężenia Ranviera M spolaryzowane, tzn. na zewnątrz błony
Wyszukiwarka
Podobne podstrony:
DSCN6607 (Kopiowanie) ■> i A Człowiek szkodliwe bakterie) i witaminy. Najbardziej nasilone zmiany
DSCN6617 (Kopiowanie) $. Człowiek $. Człowiek Ryc. 8 - 50. Akomodacja oka (opis w tekście) (JD). Oko
DSCN6625 (Kopiowanie) t. Człowiek 75 nerwowego sakomórki glejowe. Neurony pod względem budowy mogą b
DSCN6669 (Kopiowanie) S, Człowiek 119 wiążą dopełniacz i stymulują komórki NK (ang.: naturaI killers
DSCN6678 (Kopiowanie) & Człowiek 127 & Człowiek 127 węzeł zatokowy 8-88. Budowa układu bodźc
DSC00917 (2) Transport śluzowo-rzęskowy Powstająca w ten sposób fala rzęsek określana jest jako tran
154 4 koła, włożył w ten sposób szatę Nessosa. Moralna tość człowieka zaczyna się dopiero tam, gdzie
Allport - zachowanie może być niekonsekwentne, ale przez ten fakt traci cechę. Człowieka poznaje się
38979 P1090029 (2) jego aniołów. poziomie jego intencji, tj. zazdrości:,.zazdrości? mu [człowiekowi]
DSCN6540 (Kopiowanie) i zagadnienia, które do człowieka odnosić się nie będą. Ssaki to zwierzętastal
DSCN6583 (Kopiowanie) U człowieka różne tkanki odznaczają się w różnym stopniu zdolnościami do regen
więcej podobnych podstron