
Prezentacja zawiera informacje przekazane podczas wykładów
Prezentacja zawiera informacje przekazane podczas wykładów
z genetyki zwierząt dla II roku MSB
z genetyki zwierząt dla II roku MSB
rozpowszechnianie tej prezentacji
rozpowszechnianie tej prezentacji
ZABRONIONE!!
ZABRONIONE!!
Cechy jakościowe

Haploidalna liczba chromosomów- n
[2n] :
świnia
19
[38]
owca
27
[54]
bydło
30
[60]
koń
32
[64]
pies
39
[78]
kot
19
[38]
mysz
20
[40]
Kura
39
[78, w tym 60
mikrochromosomów]
Wielkość genomu ssaków
Całkowita długość DNA : około 3 mld pz
Całkowita długość genetyczna : 2500-3200
cM

Genetyka klasyczna
Cecha – właściwość organizmu łatwa do odróżnienia w zespole
innych właściwości
Genetyka klasyczna
gen = cecha
Genetyka biochemiczna
gen = łańcuch polipeptydowy
II prawo Mendla -
niezależne dziedziczenie cech
I prawo Mendla
-
prawo czystości gamet - gameta zawiera
tylko jeden allel
z danej pary alleli
Cechy
jakościowe
ilościowe
[monogenowe]
[poligenowe]

Podstawowe mechanizmy dziedziczenia cech
Współdziałanie alleliczne
Dziedziczenie z dominacją zupełną
Typ Pisum
Stosunek
fenotypów
1: 3
genotypów
1:2:1

Dziedziczenie z dominacją niezupełną – dziedziczenie pośrednie
Typ Zea
Stosunek
fenotypów
1:2:1
genotypów
1:2:1
Współdziałanie alleliczne

Kodominacja
współdominacja
Grupa krwi AB – kodominacja alleli I
A
i I
B
Współdziałanie alleliczne

Współdziałanie alleliczne
Plejotropia
Naddominacja
AA < Aa > aa
Bydło
Locus Rn – RnRn –
syndrom białych
jałówek rasy :
1. Belgijska
niebieska
2. Shorthorn
Rzekoma – szurpatość drobiu
Lisy
Locus W –
allele W i W
p
letalne
w homozygocie
W
p
W - letalny
(platynowy/białop
yski)
Właściwa – niektóre geny umaszczenia
wykazują
działanie plejotropowe
Owce karakuły
Allel W
rn
–
letalny w homo-
zygocie
Podstawowe mechanizmy dziedziczenia cech
Allele wielokrotne
Warunkują tylko jedną cechę
Każdy osobnik może mieć tylko 2 allele z serii (szeregu alleli wielokrotnych)
W populacji możne być wiele genotypów
Współdziałanie alleli z serii - dominowanie lub kodominowanie
Allele z serii można uszeregować w kolejności dominowania
Liczba genotypów zależy od liczny alleli w serii :
n (n+1) seria 13 alleli : 13 (13+1) = 91 genotypów
2
2
Układy grupowe krwi Umaszczenie Polimorficzne białka

Układy grupowe krwi niektórych
gatunków
zwierząt gospodarskich
Gatun
ek
Układy
grupow
e (loci)
Lokalizac
ja
chromos
o-
mowa
Liczba
antyge
-
nów
Liczb
a
alleli
Bydło
A
B
C
F
J
L
M
R’
S
T’
Z
15
12
18
17
11
3
23
16
21
19
10
5
40
12
5
4
1
3
2
8
1
2
10
1000
77
4
4
2
3
2
15
2
3
Allele wielokrotne

Gatun
ek
Układy
grupow
e (loci)
Lokalizac
ja
chromos
o-
mowa
Liczba
antyge
-nów
Liczb
a
alleli
Świni
e
A
B
C
D
E
F
G
H
I
J
K
L
M
N
O
1
?
?
12
9
?
15
6
18
7
9
4
11
9
6
3
2
1
2
17
4
2
5
2
2
7
12
12
3
2
2
2
2
2
15
4
2
7
2
3
6
6
18
3
2
Konie
A
C
D
K
P
Q
U
20
?
?
2
?
?
?
7
1
17
1
4
3
1
11
2
26
2
8
5
2
Układy
grupowe
krwi
niektórych
gatunków
zwierząt
gospodarskich

System
Sym-
bol
Geny
N
anty-
genó
w
Chromoso
m
ABO
Rh
MN
P
Luthera
n
Kell
Lewis
Duffy
Kidd
Diego
Cartwri
ght
Xg
Scianna
Dembro
ck
Colton
LW
Chido/
Rodgers
H
Kx
Gerbich
Cromer
Knops
Indian
ABO
RH
MNS
P1
LU
KEL
LE
FY
JK
D1
YT
XG
SC
DO
CO
LW
CH/R
G
H
XK
GE
CRO
M
KN
IN
ABO
RHD,C
E
GYPA,
B,E
P1
LU
KEL
FUT3
FY
JK
AE1
ACHE
XG
SC
DO
AQP1
LW
CH/RG
FUT1
XK
GYPC
DAF
CR1
CD44
4
45
40
1
18
23
3
6
3
7
2
1
3
5
3
3
9
1
1
7
10
5
3
9q34
1p34-
p36
4q28-
q31
22q11-
qter
19q12-
q13
7q32-
q36
19p23
1q22-
q23
18q11-
q12
17q21
7q22.1-
q22.3
Xp22-
pter
1p32-
p34
?
7p14
19p11-
p13
6p21.3
19q
Xp21.1
2q14-
q21
1q32
1q32
11p13
Systemy grupowe krwi
u ludzi
Funkcja biologiczna
systemów grupowych krwi
(5 kategorii funkcjonalnych
antygenów grup krwi)
transportery i kanały
receptory i ligandy
cząsteczki adhezyjne
enzymy
białka strukturalne

Umaszczenie zwierząt
Biochemiczne funkcje genów umaszczenia
Locus C
liczba i
intensywność
ziaren pigmentu
Locus A
rozmieszczenie
pigmentu
A – agouti a- nie
agouti
Locus B
synteza eumelaniny
Locus E
ilość produkowanej
eumelaniny
Locus D
rozjaśnienie barwy
Inne loci : W, G, Z, Sp, ...
–
różne rodzaje
umaszczenia
Allele wielokrotne

Genotypy niektórych
umaszczeń koni
Geny warunkujące umaszczenie
Fenotyp
czarne
/czerwo
ne
siw
e
biał
e
rozjaśnione
A
E
G
W
C
D
Z
Kary
Gniady
Kasztan
Palomino
Cremello
Myszaty
Siwy
Srebrny
jabłkowity
Biały
dominując
y
aa
A_
_ _
_ _
_ _
aa
_ _
aa
_ _
E_
E_
ee
ee
ee
E_
_ _
E_
_ _
gg
gg
gg
gg
gg
gg
G_
gg
_ _
ww
ww
ww
ww
ww
ww
ww
ww
W
w
CC
CC
CC
C
cr
C
C
cr
C
cr
CC
C_
CC
_ _
dd
dd
dd
dd
dd
D_
dd
dd
_ _
zz
zz
zz
zz
zz
zz
zz
Z_
_ _
_ / - dowolny allel z danej pary alleli

Podstawowe mechanizmy dziedziczenia cech
Współdziałanie niealleliczne
współdziałanie dwóch par alleli
geny addytywne (polimeryczne,
kumulatywne)
komplementarność epistaza
geny modyfikujące
Rozmieszczenie i wielkość białych plam – efekt alleli z 1 locus
łaciatość warunkowana allelami z innego locus

Komplementarność
=
dopełniające współdziałanie
genów z dwóch loci
Współdziałanie niealleliczne
Loci R i P
R
–
P
–
grzebień orzeszkowy
rr pp
grzebień pojedynczy
Rasa wyandotte
RRpp
Rasa cornish
rrPP
F
2
orzeszkowy : groszkowy
:
różyczkowy :
pojedynczy
9 : 3 : 3 : 1

Epistaza
= hamowanie
Geny z locus epistatycznego
hamują ujawnienie się genów
z locus hipostatycznego
Współdziałanie niealleliczne
Białe umaszczenie świń
epistatyczne działanie locus I

Geny addytywne (polimeryczne,
kumulatywne
)
Warunkują cechy ilościowe = produkcyjne zwierząt

Podstawowe mechanizmy dziedziczenia cech
Sprzężenie cech
Cechy dziedziczące się razem
Podczas procesu gametogenezy
geny znajdujące się w określonym
chromosomie będą przekazane
do powstającej komórki rozrodczej
łącznie, czyli cechy determinowane
przez te geny będą się dziedziczyć
razem

U muszki owocowej – w chromosomie 2
locus B - barwa ciała ( B - barwa jasnobrunatna b – czarna)
locus V - kształt skrzydeł ( V - skrzydła dłuższe od odwłoka
v – skrzydła krótkie)
Genotyp muszki dzikiej pod względem tych dwóch cech: :
BV
++
=== ===
BV
++
Genotyp podwójnej homozygoty recesywnej :
bv
===
bv
BV
Bv
faza przyciagania == faza odpychania: ===
(cis) bv
(trans) bV

Cechy sprzężone
Podstawowe mechanizmy dziedziczenia cech
Wykorzystanie sprzężenia loci w hodowli zwierząt
Mapowanie genetyczne sprzężenie między loci markerów
a loci genów „ważnych”
Analiza segregacji alleli w loci markerowych w rodzinach
referencyjnych z jednoczesnym oznaczaniem fenotypu
cech ważnych (cechy produkcyjne, zdrowotność)
Określenie odległości między loci w cM (centiMorgan)
1 cM crossing over miedzy dwoma sprzężonymi loci zachodzi
1 raz na 100 podziałów mejotycznych
Uwzględnienie trzech loci
krzyżówka trójpunktowa

Determinacja płci
[chromosomy]
1. ssaki
- płeć heterogametyczna – samce
(XY)
płeć homogametyczna –
samice (XX)
2. ptaki, niektóre owady
- płeć heterogametyczna – samice
(ZW),
płeć homogametyczna –
samce (ZZ)
3. Determinacja płci [środowisko
– temperatura]
żółwie, aligatory, niektóre
ryby
Determinacja płci
Płeć:
• chromosomowa
• gonadowa
• fenotypowa
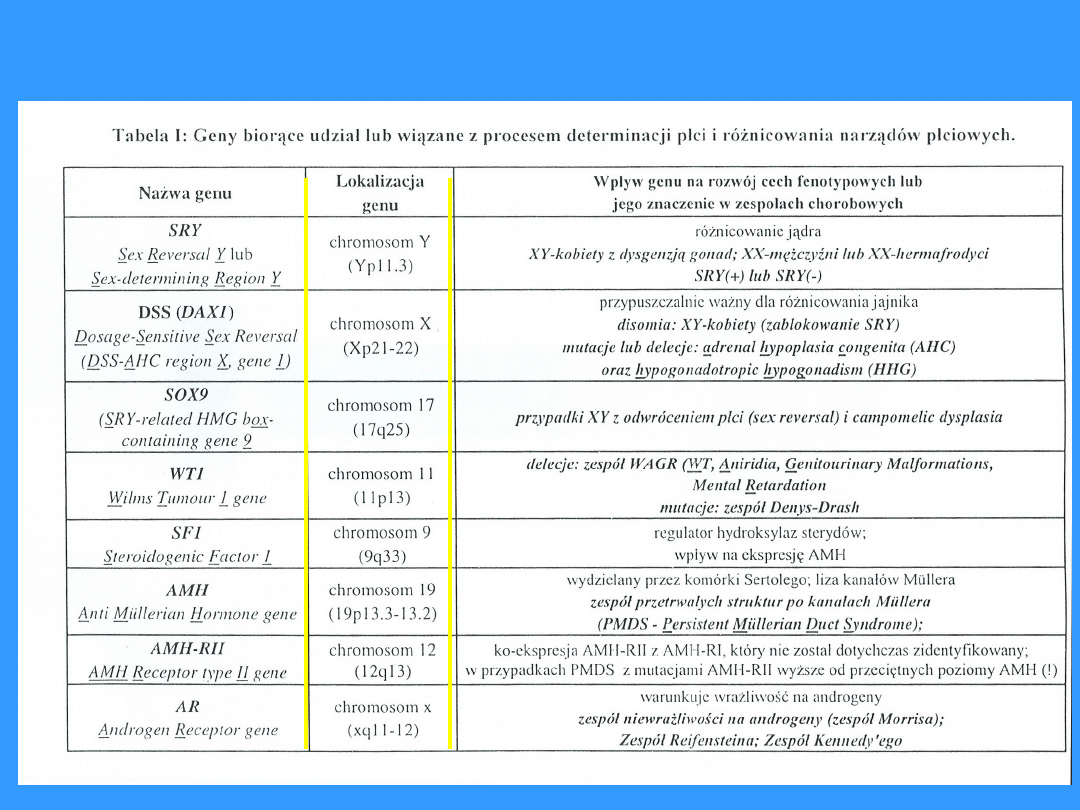
Geny warunkujące różnicowanie gonad
DMRT 1 chromosom 9 białko pełniące rolę czynnika transkrypcyjnego

WT1 – Wilm’s tumor 1 gene – koduje białko zawierające palce
cynkowe (czynnnik transkrypcyjny)
SF1 – steroidogenic factor 1 – koduje białko receptora jądrowego
regulującego ekspresję hydrolaz steroidowych
SRY
– sex determining region Y – odpowiada za tzw. przełączenie
rozowjowe – wprowadzenie niezróżnicowanej gonady
płodowej na tory prowadzące do powstania gonady męskiej
SOX9 – koduje białko pełniące rolę czynnika transkrypcyjnego
DSS
– dossage sensitive sex reversal – (DAX1) - koduje białko
receptora jądrowego dla hormonów; rozwój przewodów
Muellera a zanik przewodów Wolffa
AMH – anti-Muellerian hormone – ekspresja w komórkach Sertolego
wpływa na zatrzymanie rozwoju i zanik przewodów Muellera
DMRT1 – doublesex and Msab-3 related transcription factor 1 – ma
charakter czynnika transkrypcyjnego i pełni rolę w prawidłowym
rozwoju płci męskiej
Geny warunkujące różnicowanie gonad

Zakłócenia w determinacji płci - interseksualizm
Interseksualizm
= obojnactwo zaburzenie
procesu determinacji
i różnicowania płci wrodzone wady
rozwojowe układu
rozrodczego
Przyczyny
mutacje chromosomów płci
mutacje genów zaangażowanych w
proces
determinacji i różnicowania płci
nieprawidłowy przebieg ciąży (np.
frymartynizm)

Kategorie interseksualizmu :
hermafrodytyzm prawdziwy
= równoczesna obecność gonad
męskich i żeńskich lub struktura
złożona typu jajnikojądro
pseudohermafrodytyzm męski
= gonadom męskim towarzyszą
zaburzenia w przekształcaniu
się przewodów Wolffa i Muellera
lub powstawaniu zewnętrznych
narządów płciowych
Zakłócenia w determinacji płci - interseksualizm

frymartynizm
Zaburzenie rozwoju cech płciowych samicy z ciąży
bliźniaczej różnopłciowej
skutek powstania połączeń naczyniowych
(anastomoz) między łożyskami rozwijających się
płodów związki hormonalne oraz inne czynniki
aktywne produkowane przez jądra płodowe docierają
do organizmu samicy i wpływają na proces
różnicowania się żeńskich cech płciowych
W krwi obojga bliźniąt (samicy- frymartyna i samca)
obecność dwóch linii komórkowych – własnej i
współbliźniaka chimeryzm leukocytarny XX/XY
(badanie cytogenetyczne) i chimeryzm erytrocytarny
(badanie grup krwi)
Zakłócenia w determinacji płci - interseksualizm

Cechy sprzężone z płcią
Geny warunkujące te cechy znajdują się na chromosomie X
Płeć homogametyczna samice [ssaki] XX samce [ptaki] ZZ
Płeć heterogametyczna samce [ssaki] XY samice [ptaki] ZW
U osobnika płeci heterogametycznej - hemizygota X
A
Y [A-] Z
A
W [A-]

Inaktywacja chromosomu X w zarodkach żeńskich
Forma piętnowania podczas wczesnego rozwoju zarodkowego
Inaktywowany chromosom X podlega hetetrochromatynizacji [kondensacji]
w jadrze interfazowym, ppwstaje tzw. ciałko Barra
Rozróżnienie aktywnego i niekatywnego chromosomu X w metafazie mitotycznej
po zastosowaniu metody barwienia prążkami R (odzwierciedla tempo replikacji
poszczególnych regionów chromosomowych)
Centrum inaktywacji chromosom Xq13 gen Xist (X inactivate specific
transcript) uruchomienie inaktywacji
Losowa inaktywacja
chromosomu X
metylacja DNA
chromosomu X

Cechy sprzężone z płcią
Umaszczenie szylkretowe
u kotów – tylko samice mogą być
szylkretowe [ heterozygota Oo ]
Fenotyp Samice Samce Samice
Samce
Zdrowy
X
H
X
H
X
H
Y
lub
HH H-
Nosiciel
X
H
X
h
w
Hh
Chory X
h
X
h
X
h
Y
skrócie hh
h-
na hemofilię
Cechy sprzężone z płcią u ludzi:
• hemofilia
• dystrofia mięśniowa Duchenne’a
• dystrofia mięśniowa Beckera
• daltonizm
• rybia łuska
• albinizm [bielactwo]
• barwnikowe zwyrodnienie siatkówki
• zespół łamliwego chromosomu X
• zespół Lescha-Nyhana
• rozszczep podniebienia

Cechy sprzężone z płcią
Wykorzystanie w hodowli zwierząt
Rasy drobiu autoseksingowe barwa puchu
marker płci
Rasa Polbar gen jastrzębiatości wyhodowana w Polsce
w połowie XX w. przez Laurę Kaufman

Zjawisko rodzicielskiego piętnowania genomu
obswerwowane u wyższych
Eucaryota polega na naznaczeniu chromosomów
zgodnie z ich
rodzicielskim pochodzeniem i prowadzi do
zróżnicowanej ekspresji alleli
ojcowskich i matczynych podczas rozwoju osobniczego
Piętno genomowe = piętno rodzicielskie = piętno
gametyczne (ang. gametic imprinting)

Piętno rodzicielskie
Metylacja DNA przyłączenie grup
metylowych powoduje przejście
DNA w stan nieaktywny (wskutek
reorganizacji nici nukleosomowej)
oraz zmianę powinowactwa DNA
do czynników transkrypcyjnych
nie wiadomo czy metylacja
poprzedza czy następuje
po
inaktywacji genów
sprzężonych
z chromosomem X
genom plemników jest
bardziej
metylowany niż genom
komórki
jajowej

Piętno rodzicielskie
przykład z genetyki człowieka
Zespoły Pradera-Willego i Angelmana spowodowane są
utratą funkcji
odrębnych, sprzężonych genów, które poddane są
piętnowaniu
rodzicielskiemu
Delecja fragmentu przycentromerowego ramienia q
chromosomu 15 :
Uszkodzony chromosom pochodzi od ojca – zespól
Pradera-Willego
(objawy m.in. otyłość i niedorozwój umysłowy)
Uszkodzony chromosom pochodzi od matki – zespół
Angelmana
(objawy m.in. głębokie upośledzenie umysłowe, brak
mowy, drgawki)
Region 15q11-q13
• gen UBE3A (ubiquitin protein ligase E3) - nie podlega
ekspresji
jeśli pochodzi od ojca
• kilka genów (m.in. SNRPN – small nuclear
ribonucleoprotein
polypeptide N) - nie podlega ekspresji jeśli pochodzi
od matki

Zespół Angelmana (AS)
- częstość 1/10 000-20
000 urodzeń
Zespół Angelmana może być spowodowany brakiem
regionu
15q11q13 na chromosomie pochodzącym od
matki lub mutacją
w genie UBE3A lub disomią ojcowską (oba
chromosomy 15
pochodzą od ojca)
Zespół Pradera-Willego (PWS)
- częstość 1/10 000-20 000 urodzeń
W zespole Pradera-Willego defekt molekularny polega na braku
regionu 15q11q13 w chromosomie pochodzącym od ojca
lub disomii matczynej
W zygocie i potomnych pokoleniach komórek
somatycznych stan
napiętnowania genu (-ów) jest utrzymywany –
odstępstwa :
• gen napiętnowany podczas gametogenezy
powtórnie piętnowany
podczas rozwoju osobniczego - gen Ifg2 (insulin
growth factor 2)
u myszy
Piętno rodzicielskie

DNA mitochondrialny
Organizm
Liczba par
zasad
____________________________
Ssaki
człowiek
16 569
bydło
16
338
owca
16580
mysz
16
295
wieloryb
16 398
Ptaki
kura
16
775
Ryby
trzonopłetwe
16
407
Stawonogi
muszka owocowa
16
019
Nicienie
jelitowe świń
14
284
nicienie glebowe13 794
Wielkość (w parach zasad) mtDNA

DNA mitochondrialny
Kompleks
I
II
III
IV
V
Suma
mtDNA
7
0
1
3
2
13
jądrowy DNA
33 4
10
10
10
67
Większość komórek zawiera od tysiąca do 10 tys. kopii
mtDNA
Jedynie w oocytach II rzędu liczba kopii jest wyraźnie
większa i sięga
około 100 tys.
Prawie cały mtDNA zawiera sekwencje kodujące, a
sekwencje
powtarzające się tandemowo są reprezentowane bardzo
nielicznie
Łańcuch ciężki 12 genów kodujących białka, 14 genów
kodujących tRNA
i 2 geny kodujące rRNA
Łańcuch lekki tylko jeden gen kodujący białko i 8 genów
kodujących
cząsteczki tRNA

mtDNA
jądrowy DNA
Podwójna nić kolista
Podwójna nić spiralnie
zwinięta
Brak intronów
Obecność
intronów
Transkrybowane obie nici
Transkrybowana
jedna nić
Replikacja jednoczesna
Replikacja
niejednoczesna
DNA „nagi” bez białek DNA
powiązany z białkami
DNA mitochondrialny
Document Outline
- Slide 1
- Slide 2
- Slide 3
- Slide 4
- Slide 5
- Slide 6
- Slide 7
- Slide 8
- Slide 9
- Slide 10
- Slide 11
- Slide 12
- Slide 13
- Slide 14
- Slide 15
- Slide 16
- Slide 17
- Slide 18
- Slide 19
- Slide 20
- Slide 21
- Slide 22
- Slide 23
- Slide 24
- Slide 25
- Slide 26
- Slide 27
- Slide 28
- Slide 29
- Slide 30
- Slide 31
- Slide 32
- Slide 33
- Slide 34
- Slide 35
- Slide 36
- Slide 37
Wyszukiwarka
Podobne podstrony:
studenci2 cechy jakościowe
studenci cechy jakościowe
studenci cechy ilościowe
studenci cechy ilociowe
studenci2 cechy jakościowe
7a interpretacje i cechy info, Procesy informacyjne w zarządzaniu, materiały student Z-sem 12-13, wy
Statystyka opisowa - rozkład jednej cechy - powtórkowe, Przydatne Studentom, Akademia Ekonomiczna Kr
1 cechy fizyczne gruntow dla studentow
Cechy dobrego nauczyciela student
wyklad 1 cechy fukcje regulacje instrumenty dla studentow
2010 ZMP studenci
gruźlica dla studentów2
Prezentacja 2 analiza akcji zadania dla studentow
Szkolenie BHP Nowa studenci
Student Geneza
Kosci, kregoslup 28[1][1][1] 10 06 dla studentow
więcej podobnych podstron