1. EWOLUCJA PREBIOTYCZNA.
Prekursory replikatora. Chromosomy zostaly odkryte przez Waldeyera juz ponad 100 lat temu, w 1888r., jako intensywnie barwiace sie (stad nazwa) organelle, ktore pojawiaja sie w komorce eukariotow tuz przed podzialem mitotycznym i ktore zanikaja w powstajacych jadrach komorkowych po jego zakonczeniu. W 1902 r. Sutton wysunal hipoteze, ze chromosom jest nosicielem mendlowskich genow, co rozwoj biologii molekularnej potwierdzil. Jednak dopiero obecnie zaczynamy zdawac sobie sprawe z konsekwencji jakie nawet niepelna wiedza o strukturze chromosomu i technologie wypracowane w trakcie jej zdobywania pociagaja za soba, jak zaczynaja wplywac na nasze zycie a takze na sposob, w jaki patrzymy na samych siebie i na otaczajacy nas swiat.
Obecnie definiuje sie chromosom jako nosnik cech dziedzicznych organizmow zywych oraz wirusow, rodzaj bazy danych, zbudowanej z kompleksu kwasow nukleinowych (najczesciej DNA) z bialkami. Suma informacji genetycznej zawartej w chromosomach jakiegos gatunku (wirusa, rosliny, bakterii czy zwierzecia) lub w DNA mitochondrialnym czy chloroplastycznym, nazywa sie genomem i moze byc zapisana w postaci sekwencji nukleotydowej czasteczki DNA, ale mozna ja zapisac rowniez na kartce papieru, na dysku kompaktowym czy w pamieci komputera.
Okolicznosci zwiazane z pojawieniem sie pierwszego chromosomu sa przedmiotem licznych spekulacji m.in. rowniez tej, ze zostal zawleczony na ziemie celowo lub przypadkiem. Nawet jesli zdarzenie takie rzeczywiscie mialo miejsce, to hipoteza ta nie odpowiada na pytanie skad sie wzial pierwszy chromosom. Wsrod specjalistow przewaza opinia, ze przodek wszystkich chromosomow, z ludzkimi wlacznie, byl produktem miejscowym.
Warunki jakie panowaly na ziemi przez pierwsze 500 milionow lat po powstaniu ukladu slonecznego, nieco ponad 4.5 miliarda lat temu, sprzyjaly raczej rozkladowi zwiazkow organicznych niz ich syntezie. Z drugiej strony, pierwsze geologiczne formacje, ktorych sklad chemiczny i izotopowy nie daje sie wytlumaczyc inaczej jak tylko zakladajac istnienie organizmow zywych (a wiec posiadajacych chromosomy) datuje sie na okolo 3.85 miliarda lat. Co dzialo sie w okresie kilkuset milionow lat, podczas ktorych te organizmy sie pojawily wlasciwie nie wiadomo (ryc. 1 - 1). Wiekszosc badaczy sadzi, ze atmosfera ziemska w tym okresie byla beztlenowa i zawierala glownie gazy takie jak azot, wodor, metan, dwutlenek wegla, cjanowodor, amoniak i para wodna. Doswiadczenia Millera (ryc. 1 - 2) pokazaly, ze wyladowania elektryczne w mieszaninie zawierajacej metan, amoniak, wodor i pare wodna powoduja powstanie wielu aminokwasow w tym rowniez takich, ktore wystepuja w organizmach zywych. Inni, zmieniajac nieco warunki, udowodnili powstawanie adeniny, cytozyny i uracylu a takze cukrow, w tym i rybozy, z takich prostych skladnikow jak para wodna, azot, metan, cjanowodor i formaldehyd (ryc. 1 - 3). Jesli w atmosferze ziemskiej rzeczywiscie panowaly kiedys warunki zblizone do tych, jakie stosowali autorzy wzmiankowanych doswiadczen, to niewatpliwie w kaluzach, jeziorach i oceanach mlodej podowczas ziemi rozpuszczone byly zwiazki chemiczne stanowiace elementarne cegielki struktury chromosomu. Ten hipotetyczny roztwor nazywany jest bulionem pierwotnym lub zupa prebiotyczna.
Chyba kazdy z nas wykonywal w ramach lekcji fizyki doswiadczenie polegajace na tym, ze nasycony roztwor np. sacharozy, w ktorym zawieszono na nitce krysztalek cukru, pozostawiano do powolnego odparowania. Po kilku dniach stwierdzic mozna bylo, ze krysztal powiekszyl sie zachowujac jednak forme szescianu. Rosnacy krysztal niejako wymusza takie przestrzenne usytuowanie sie kolejnych adsorbujacych sie na jego powierzchni czasteczek, ze geometria sieci zostaje zachowana. Na podobnej zasadzie powstaly prawdopodobnie pierwsze czasteczki polimeru, ktore moznaby nazwac proto-chromosomem. Wykazano m.in. ze siec krystaliczna pospolitego, wystepujacego w glinie mineralu montmorylonitu, ulatwia ustawianie sie w szereg i polimeryzacje aktywowanego rybonukleotydu (5'fosforimidazolidu adenozyny) w wyniku czego powstaje oligomer RNA (ryc. 1 - 4). Doswiadczenia tego typu maja na celu, przynajmniej na razie, nie tyle odtworzenie warunkow w ktorych powstal pierwszy chromosom, ile udowodnienie, ze spontaniczna polimeryzacja nukleotydow, ktora warunkuje powstanie jego prymitywnej formy, tzw. replikatora, jest w ogole mozliwa. Replikator jest to hipotetyczna czasteczka lub kompleks czasteczkowy, ktory ma zdolnosc tworzenia swojej wlasnej kopii, badz bezposrednio, badz tez syntetyzujac uprzednio matryce na ktorej ta kopia powstaje. Ma wiec zdolnosc rozmnazania sie. Kolejne kopie replikatora, ktory kiedys pojawil sie w zupie prebiotycznej roznily sie od siebie w drobnych szczegolach struktury a co za tym idzie w trwalosci i we wlasnosciach katalitycznych. Najtrwalsze z replikatorow z definicji trwaly dluzej niz inne a najlepsze autokatalizatory najszybciej sie rozmnazaly. Powstawaly pewnie i takie replikatory, ktore rozkladaly inne replikatory na elementy skladowe i z tych elementow budowaly wlasna strukture. Replikator, ktory mial sklonnosci do izolowania sie od otoczenia utworzyl rodzaj prymitywnej komorki.
Jesli chodzi o nature chemiczna pierwszych replikatorow jestesmy skazani na domysly. Prawdopodobnie nie byly to czasteczki kwasu dezoksyrybonukleinowego i niekoniecznie byly to oligorybonukleotydy (ryc. 1 - 5). Jakkolwiek by nie bylo, i replikatory i sklad prebiotycznego bulionu ewoluowaly. Przypuszcza sie, ze po pewnym okresie powstalo w nim cos co sie okresla nazwa « swiat RNA ». Kompleksy makromolekularne istniejace w tym swiecie mialy « chromosomy » zbudowane z kwasu rybonukleinowego. Z RNA zbudowane byly rowniez katalizatory (rybozymy) ktore umozliwialy zachodzenie reakcji niezbednych dla trwania i rozmnazania sie tych kompleksow. Nie istnial wiec podzial na fenotyp i genotyp ani potrzeba translacji. Zsyntetyzowano w laboratorium szereg rybozymow bedacych w stanie katalizowac rozne reakcje biochemiczne, co czyni istnienie « swiata RNA » prawdopodobnym. Za tym, ze « swiat RNA » rzeczywiscie istnial przemawia rowniez fakt, ze niektore wspolczesne organizmy stosuja rybozymy mimo, ze bialka potrafilyby z pewnoscia wykonywac podobne funkcje rownie dobrze, jesli nie lepiej. Sadzi sie, ze wlasnie rybozymy stanowia relikty tego zaginionego swiata. Co prawda nie zsyntetyzowano jeszcze replikatora, ktory mialby oczekiwane wlasnosci, ale mozna sadzic, ze jest to tylko kwestia czasu i ze z mala pomoca uczonych synteza ta potrwa krocej niz za pierwszym razem.
Wlasnosci katalityczne RNA poznano przy okazji badan nad samowycinajacymi sie intronami.
Introny. Przewazajaca czesc intronow obecnych w eukariotycznym pre- mRNA to introny typu GU-AG (nazwane tak poniewaz te dinukleotydy znajduja sie po stronie 5' i 3' wycinanego segmentu RNA. W zasadzie te dinukleotydy sa czescia dluzszych sekwencji typu konsensus. Po stronie 5' intronu znajduje sie sekwencja 5'AGGUAAGU 3' a po stronie 3' sekwencja 5'PyPyPyPyPyPyNCAG3' (Py oznacza C lub U) (ryc. 1 - 6). Takie introny ulegaja wycinaniu przy pomocy bialkowych systemow enzymatycznych. Podobne wlasciwosci maja rzadkie introny AU-AC. Niewielka ilosc takich intronow znaleziono u drosofili u czlowieka i u roslin.
Wyciecie intronu GU-AG zaczyna sie od ataku grupy hydroksylowej 2' adenozyny znajdujacej sie wewnatrz intronu, ktora tworzy wiazanie z fosforanem 5' guanozyny w dinukleotydzie GU tworzac strukture typu « lasso ». Uwolniona grupa 3'OH eksonu atakuje wiazanie po drugiej stronie intronu eliminujac go z produktu transkrypcji. Wycinanie intronu jest reakcja prosta biochemicznie. Trudnosc polega na tym, ze eksony moga byc oddalone jeden od drugiego o kilkadziesiat kb (100 nm i wiecej). System wycinajacy musi znalezc to poprawne miejsce. Poniewaz sekwencje flankujace eksony sa podobne a geny maja ich niekiedy po kilkadziesiat, istnieje mozliwosc opuszczenia jednego czy kilku eksonow. Ponadto wewnatrz intronow istnieja miejsca o sekwencji podobnej do sekwencji rozpoznawanej przez aparat wycinajacy. Latwo wiec o pomylke. Aparat wycinajacy introny sklada sie z szeregu bialek oraz z snRNA (sn - small nuclear) U1 - U6 . Zawieraja one od 106 (U6) do 185 (U2) nukleotydow. W wycinaniu intronow AU-AC biora udzial snRNA U5 oraz U4atac, U6atac, U11 i U12. Z punktu widzenia ewolucji najbardziej interesujace sa introny, ktore nie potrzebuja bialek poniewaz badajac je wlasnie zdano sobie sprawe, ze RNA moze miec wlasnosci katalityczne.
Przykladem samowycinajacego sie intronu jest intron obecny w genie kodujacym rRNA u tetrahymena. Nalezy on do tak zwanej grupy I intronow. Odnaleziono je rowniez u bakteriofaga T4, w genomach mitochondriow, chloroplastow a takze u bakterii. Reakcja transestryfikacji jest podobna do reakcji intronow pre-mRNA, ale wymaga kofaktora jakim jest guanozyno - mono-, dwu- lub troj- fosforan. 3'OH tego kofaktora atakuje wiazanie fosfodwuestrowe od strony 5' miejsca wycinania i wiaze sie z koncem 5' intronu. Druga transestryfikacja odbywa sie z udzialem 3'OH na koncu eksonu, ktore to OH atakuje wiazanie fosfodwuestrowe po stronie 3' w miejscu wycinania intronu. W wyniku tego ataku eksony ulegaja polaczeniu a intron wycieciu (ryc 1 - 7). Produkt reakcji jest w tym przypadku liniowy, chociaz pozniej tworzy czasteczke kolowa i ulega degradacji. W organellach i u niektorych prokariotow znaleziono rowniez innego typu introny, tzw. grupy II i III rozniace sie struktura od intronow grupy I. Samowycinajace sie introny maja skomplikowana strukture drugo i trzeciorzedowa, co jest niezbedne do tego aby mogly one przejawiac wlasnosci katalityczne (ryc. 1 - 8) i roznia sie tym od innych intronow, takich np. jak introny grupy czwartej znajdujace sie w eukariotycznych t-RNA. Ich wycinanie nie wymaga transestryfikacji tylko dzialania rybonukleazy, ktora zostawia cykliczny fosforan w miejscu 3' i grupe OH w miejscu 5'. Te konce wiaza sie z udzialem ligazy RNA.
Synteza i ewolucja ligazy in vitro. Jeden z intronow pochodzacych z genu sunY z faga T4 zostal uzyty do skonstruowania rybozymu, ktory katalizowal reakcje ligacji (ryc. 1 - 9). W pierwszym etapie zmodyfikowano sekwencje tego intronu przez delecje koncow 3' i 5' (w sumie okolo 40 nukleotydow). Nastepnie zastapiono kilka nukleotydow innymi, obecnymi w homologicznych miejscach w genie td dla zapewnienia wiekszej stabilnosci struktury przestrzennej czasteczki. Jako substratu reakcji ligacji, ktora ten zmodyfikowany intron mial katalizowac uzyto dwoch oligonukleotydow tworzacych strukture dwuniciowa (ryc. 1 - 10). Ligacja odbywala sie z uwolnieniem guanozyny. Przebieg reakcji sledzono obserwujac powstawanie wysokoczasteczkowego produktu z substratu o nizszym ciezarze czasteczkowym. W dalszym etapie doswiadczenia autorzy przecieli petle P6 i P8 w rybozymie w wyniku czego otrzymali oligonukleotydy A, B i C o dlugosci 59, 75 i 83 nukleotydow (ryc. 1 - 11). Powstaly trojskladnikowy kompleks mial nieco nizsza optymalna temperature reakcji i nieco nizsza aktywnosc katalityczna. Potrafil on jednak katalizowac autoligacje prowadzaca do powstania jednej czasteczki z mieszaniny oligorybonukleotydow. Jak wynika z tego synteza pierwszych replikaz nie musiala byc tak trudna jak to mogloby sie wydawac.
Sukces ten zachecil badaczy do podjecia prob otrzymania ligazy calkowicie syntetycznej w drodze selekcji in vitro uzywajac jako produktu wyjsciowego mieszaniny syntetycznych oligonukleotydow posiadajacych przypadkowa sekwencje nukleotydowa. Schemat doswiadczenia jest przedstawiony na ryc. 1 - 12. Procedure tego typu stosuje sie obecnie dosc szeroko. Nazywa sie ja SELEX (systematic evolution of ligands by exponential enrichment) do otrzymywania aptamerow (aptus lac. pasujacy) interesujacych z farmakologicznego punktu widzenia.
Aby otrzymac syntetyczny rybozym posiadajacy aktywnosc ligazy RNA (Ryc 1-12A) pierwszym etapie zsyntetyzowano mieszanine oligonukleotydow o przypadkowych sekwencjach z ktorych kazdy mial dinukleotyd pppGG na koncu 5' oraz sekwencje stala, komplementarna do startera odwrotnej transkryptazy, na koncu 3' (patrz nizej). Te mieszanine inkubowano z substratem, ktorym byla czasteczka biotynylowanego oligonukleotydu RNA posiadajaca sekwencje rozpoznawana przez enzym restrykcyjny Bcg I [5'…↓(N)10CGA(N)6TGC(N)12↓…3']. Te z czasteczek, o sekwencji przypadkowej, ktore mialy wlasnosci katalityczne ligazy wiazaly wiazaniem fosfodwuestrowym swoj koniec 5' z koncem 3' substratu z uwolnieniem pirofosforanu. Produkt reakcji byl izolowany z mieszaniny substratow stosujac kuleczki magnetyczne pokryte streptawidyna - bialkiem posiadajacym powinowactwo do biotyny. Oddzielony produkt zostal nastepnie uzyty do otrzymania komplementarnego DNA w reakcji z odwrotna transkryptaza. RNA matrycy zostal wytrawiony a otrzymany jednoniciowy DNA zostal namnozony w reakcji PCR przy uzyciu dwoch starterow. Jeden z nich byl komplementarny do sekwencji stalej obecnej na koncu sekwencji przypadkowej a drugi mial sekwencje taka jak koniec 5' substratu. Tak uzyskany DNA trawiono enzymem Bcg I zostawiajacym czasteczki DNA zakonczone dinukleotydem 3'-CC poniewaz dwa ostatnie nukleotydy w sekwencji rozpoznawanej przez enzym Bcg I (patrz powyzej) byly guaninami. Do powstalego oligonukleotydu ligowano promotor T7. W wyniku transkrypcji, ktora potem przeprowadzano otrzymywano czasteczki RNA posiadajace juz pewna aktywnosc katalityczna ligazy. Czasteczki te uzywano w drugim cyklu selekcji. Po przeprowadzeniu kilku takich cykli otrzymano mieszanine rybozymow majacych zdolnosc katalizowania reakcji ligacji RNA. Rycina 1 - 13 przedstawia kompleks jednego z takich syntetycznych rybozymow z substratem. Zdolnosc tego rybozymu do katalizowania reakcji ligacji pozwalala przypuszczac, ze moze on rowniez katalizowac reakcje polimeryzacji aktywowanych nukleotydow na matrycy RNA. I rzeczywiscie, stwierdzono, ze syntetyczny rybozym dodaje trzy komplementarne nukleotydy do substratu jesli substrat ten tworzy odpowiedni kompleks z rybozymem (ryc. 1 - 14).
Synteza i selekcja in vitro rybozymu katalizujacego rybozylacje zasady purynowej. Uwaza sie, ze nukleotydy byly rzadkim skladnikiem bulionu pierwotnego. Istotnym etapem ewolucji musiala byc wiec synteza rybozymu zdatnego do syntezy nukleotydow z prekursorow takich jak zasady azotowe i fosforany rybozy. Unrau i Bartel przedstawili system selekcji in vitro przy zastosowaniu ktorego otrzymali polirybonukleotyd zdolny do katalizowania takiej reakcji. Reakcja, ktora rybozym katalizowal byla to synteza 5' fosforanu urydyny z uracylu i fosfopirofosforanu rybozy (pRpp). We wspolczesnych organizmach reakcja taka jest katalizowana przez fosforybozylotransferaze uracylowa (UPRT, ryc. 1 - 15). Jest ona kluczowym elementem lancucha reakcji prowadzacych do biosyntezy nukleotydow. Nie jest znany rybozym naturalny, ktory bylby zdolny katalizowac te reakcje. Postanowiono sprawdzic czy droga selekcji in vitro mozna taki rybozym wyodrebnic z mieszaniny polinukleotydow RNA posiadajacych przypadkowa sekwencje oligorybonukleotydow. Rycina 1 - 16 pokazuje w jaki sposob rybozym taki zostal wyselekcjonowany.
Najpierw autorzy zsyntetyzowali pule oligonukleotydow typu 5'-(promotor-T7)-(17-Nt-constant)-N76-CCTG-N76-GGCACC-N76-(17-Nt-constant)-3' posiadajacych 228 pozycji zawierajacych przypadkowe nukleotydy. Ogolem do selekcji wzieto mieszanine zawierajaca 1.5x1015 roznych czasteczek RNA. Na poczatku kazdego cyklu selekcji pRpp byl przylaczany do tych oligonukleotydow do konca 3' w reakcji z AppRpp katalizowanej przez T4 RNA ligaze. Mieszanine produktow reakcji inkubowano z analogiem uracylu, 4 tiouracylem (4SUra) aby umozliwic sekwencjom, ktore byly zdolne do tworzenia wiazania glikozydowego zwiazanie rybozy do tiouracylu z uwolnieniem pirofosforanu. RNA przylaczony do tiouracylu byl oddzielany od nieprzereagowanych substratow na zelu zawierajacym octan N-akryloiloaminofenylorteciowy. Produkt reakcji wedrowal znacznie wolniej dzieki powinowactwu siarki do rteci. Byl on nastepnie eluowany z zelu i po powtornym oczyszczeniu przez elektroforeze, biotynylowany w reakcji z jodoacetylo-LC-biotyna. RNA byl nastepnie wychwytywany przez magnetyczne kuleczki pokryte streptawidyna, co pozwalalo oczyscic RNA majacy wlasnosci katalityczne. Taki RNA byl uzyty do syntezy komplementarnego DNA w reakcji z odwrotna transkryptaza, ktory to DNA byl nastepnie amplifikowany w reakcji z PCR. Otrzymany DNA transkrybowano w celu uzyskania RNA, ktory poddano kolejnej rundzie selekcji. Po 11 cyklach aktywnosc katalityczna tak uzyskanego rybozymu byla 5x106 wyzsza niz w kontroli. Po sklonowaniu i oznaczeniu sekwencji okazalo sie, ze otrzymano trzy rodziny rybozymow zdolnych do katalizowania syntezy rybonukleotydu z 4SUra w sposob bardzo specyficzny. Reakcja wymagala Mg++ lub Mn++.
Synteza i selekcja in vitro rybozymu katalizujacego synteze wiazania peptydowego. Hipoteza swiata RNA proponuje, ze "zycie" bazujace na rybozymach musialo kiedys "nauczyc sie" syntetyzowac polipeptydy. Aby to bylo mozliwe RNA musi byc zdolne do katalizowania reakcji tworzenia wiazania amidowego uzywajac jako donora aminoacylo-RNA. Metoda selekcji in vitro otrzymano rybozym, ktory ma aktywnosc aminoacylotransferazy i ktory przenosi aminokwas z aminoacylo-RNA na samego siebie. Lohse i Szostak, ktorzy otrzymali ten rybozym sygeruja, ze reakcja ta jest analogiczna do transferu peptydylu na rybosomie. Ryc. 1 - 17 przedstawia reakcje transferu formylometioniny z fMet-t-RNA na hydroksypuromycyne (ale podobna reakcja ma miejsce gdy grupa -OH jest zastapiona przez grupe NH2 w puromycynie). Analogiczna reakcja zostala zastosowana jako podstawa selekcji in vitro rybozymu (ryc 1 - 18). Mieszanina okolo 1015 roznych czasteczek RNA zostala otrzymana przez transkrypcje syntetycznej matrycy DNA. To RNA zostalo potraktowane fosfataza, tak aby otrzymac wolny koniec 5'OH. Substratem byl 5' pCAACCAO 3'(N-biotynylo-Met), ktory nasladuje 3' fragment t-RNA. Po inkubacji substratu z RNA te sekwencje, ktore byly w stanie zwiazac biotynylowany aminokwas zostaly oddzielone od pozostalych skladnikow mieszaniny na streptawidynoagarozie. Oddzielone RNA zostaly namnozone w podobny sposob jak poprzednio tzn. przez serie reakcji retrotranskrypcji i PCR powstalego DNA, korzystajac z tego, ze flankujace segmenty RNA byly stale. Ta druga pula RNA zostala nastepnie defosforylowana i przereagowana z tym samym substratem. Pomiedzy cyklami 6 i 7 przeprowadzono mutagenizujacy PCR tak aby wzbogacic ilosc potencjalnie aktywnych sekwencji. Po jedenastu cyklach aktywnosc acetylotransferazy bylo 104 razy wyzsza niz aktywnosc mieszaniny wyjsciowej (ryc. 1 - 19). Otrzymany RNA sklonowano i zsekwencjonowano. Okazalo sie, ze otrzymane rybozymy nalezaly do trzech klas. Jedna z tych klas zawierala 13-to nukleotydowy segment niezmienny, ktorego 6 nukleotydow bylo komplementarne do sekwencji aminoacylowanego oligonukleotydu a 7 do stalego konca 5' rybozymu. Ta 13 nukleotydowa sekwencja dziala prawdopodobnie jako matryca, ktora umozliwia kontakt grupy aminoacylowej z koncem OH rybozymu.
Zsyntetyzowano nastepnie rybozym z grupa NH2 przy 5'G poprzez przeprowadzenie transkrypcji in vitro w obecnosci 5' amino-dezoksyguanozyny. W optymalnych warunkach 70% transkryptu rozpoczynalo sie od grupy 5' aminowej, podczas gdy reszta zaczynala sie od 5' trojfosforanu i nie partycypowala w reakcji. Ten zmodyfikowany rybozym katalizowal transfer aminokwasu na grupe NH2 z taka sama efektywnoscia jak na grupe -OH. Z doswiadczen tych wynika, ze RNA potrafi katalizowac synteze wiazania -CONH- rownie dobrze jak rybosomalna peptydylotransferaza.
Sprzezenie replikacji z transkrypcja. Etapem posrednim pomiedzy « swiatem RNA » a obecnym swiatem istot zywych byl prawdopodobnie « swiat » w ktorym odegraly role czasteczki typu aminoacylo-t-RNA. Jak sugerowal Gordon, na tym etapie ewolucji chemicznej mogl powstac przodek wspolczesnego rybosomu, ktory katalizowal rownoczesnie translacje i replikacje. Hipotetyczny ten proto-rybosom zawieralby rybozym ktory syntetyzujac lancuch polipeptydowy rownoczesnie wycinalby triplety antykodonow z przodkow dzisiejszych aminoacylo-t-RNA laczac je w nic matrycy RNA, komplementarnej do nici kodujacej (ryc. 1 - 20). Rybozym, ktorego istnienie postuluje Gordon mialby wiec wlasnosci zblizone do wspolczesnych intronow z grupy I. Taki intron wycina sam siebie z prekursorowego polirybonukleotydu rownoczesnie ligujac nukleotyd lezacy po swojej stronie 5' z nukleotydem lezacym po stronie 3'. Hipoteza Gordona tlumaczy jak w swiecie RNA pojawic sie mogly polipeptydy, ktore maja zdolnosc tworzenia bardziej wydajnych i trwalszych katalizatorow niz rybozymy
Dalszym etapem ewolucji bylo oddzielenie sie fenotypu od genotypu. Replikator, ktory potrafil magazynowac swoja informacje genetyczna w czasteczkach trwalego kwasu dezoksyrybonukleinowego mial przewage nad organizmami, obecnymi w swiecie RNA.
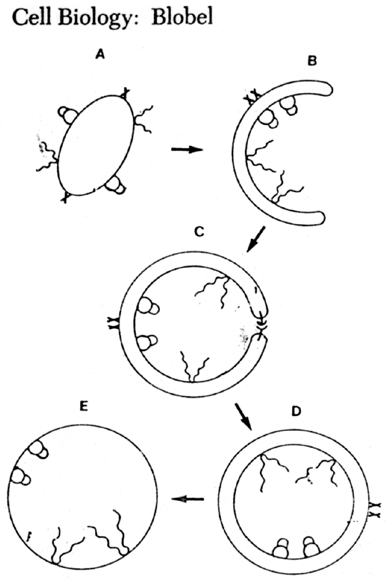
Kolejnym etapem bylo pojawienie sie pierwszej komorki (ryc. 1 - 21). W laboratorium Szostaka badano ten proces na modelu doswiadczalnym. Stwierdzono, ze micelle kwasow tluszczowych w obecnosci montmorylonitu przeksztalcaja sie w rodzaj pecherzyka w ktorym zostaja zamkniete czastki tego mineralu. Jesli montmorylonit zawiera zaadsorbowany RNA, to rowniez i on zostaje zamkniety wewnatrz pecherzyka. Jesli do zawiesiny wprowadzi sie dodatkowe micelle, pecherzyki "rosna", natomiast filtracja pod cisnieniem powoduje ich podzial. Doswiadczenia tego typu pokazuja, ze pierwotne komorki mogly tworzyc sie, rosnac i dzielic w obecnosci katalizatorow mineralnych izolujac od otoczenia i ochraniajac przed rozkladem pirwotne oligonukleotydy.
Ukoronowaniem procesow ewolucji prebiotycznej bylo pojawienie sie LUCA (Last Universal Common Ancestor) - organizmu, ktorego istnienia dowodzi biochemiczna jednosc swiata zywego, i ktory dal poczatek zarowno eukaryotom jak i prokariotom.
Na LUCA (zwanego inaczej progenota) konczy sie prehistoria i zaczyna sie «pisana» historia chromosomu. Z okresu prehistorycznego pozostaly tylko nieliczne relikty, takie jak rybozymy natomiast po dawno zaginionym LUCA przetrwaly kopie (coprawda niezbyt wierne) zapisu informacji zawartej w jego chromosomie. Kopie te to informacja zawarta w chromosomach dzisiaj istniejacych organizmow z naszym wlasnym wlacznie.
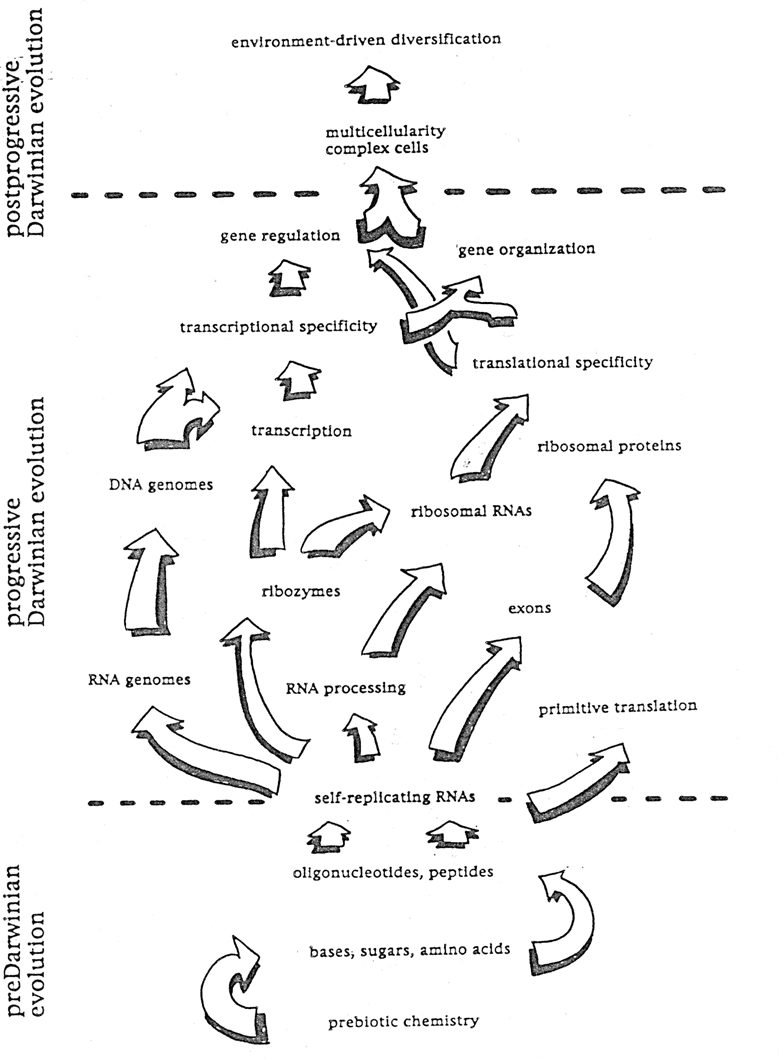
Ryc. 1 - 1 Podstawowe etapy ewolucji prebiotycznej
Ryc. 1 - 2. Aparat Millera w ktorym przeprowadzano synteze aminokwasow
Ryc. 1 - 3. Synteza prekursorow biopolimerow.
Ryc. 1 - 4. Tworzenie wiazania fosfodwuestrowego prowadzacego do syntezy 3' - 5' oligorybonukleotydow katalizowane przez adsorpcje aktywowanych monomerow na powierzchni montmorylonitu. A - reszta adeniny, R - reszta rybozy, w ramce 5' fosforimidazolid adenozyny. Strzalki pomiedzy resztami adeniny symbolizuja wiazania o charakterze niejonowym.
Ryc. 1 - 5. Biopolimery, ktore potencjalnie mogly byc nosnikami informacji genetycznej w bulionie pierwotnym.
Ryc. 1 - 6. Introny typu GU - AG
Ryc. 1 - 7. Wycinanie intronu grupy I i II
Ryc. 1 - 8. Intron w rRNA Tetrahymena.
Ryc. 1 - 9. Intron obecny w genie sunY faga T
Ryc. 1 - 10. Rybozymatyczna reakcja ligacji RNA
Ryc. 1 - 11. Kompleks oligorybonukleotydow majacy wlasciwosci ligazy.
Ryc 12. Typowa struktura aptameru i schemat metody SELEX (RT - odwrotna transkryptaza, T7 promoter- sekwencja promotorowa bakteriofaga T7)
Ryc. 1 - 12A. Synteza i selekcja in vitro rybozymu posiadajacego wlasnosci ligazy RNA. B - biotyna, ST strepawidyna,, RT transkryptaza odwrotna.
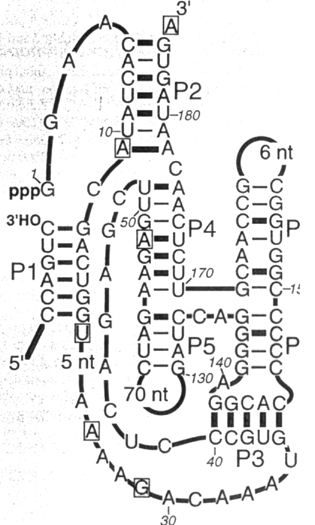
Ryc. 1 - 13. Kompleks syntetycznego rybozymu, majacego aktywnosc ligazy, z substratem.
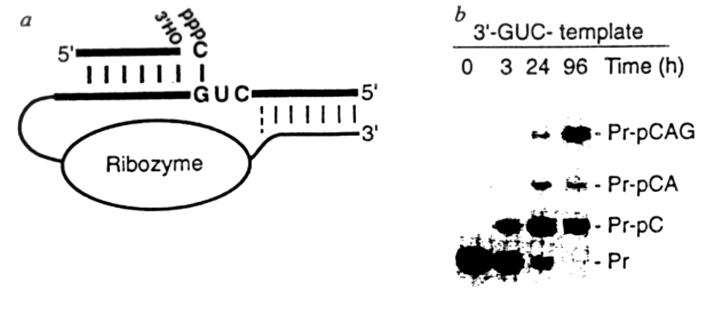
Ryc. 1 - 14. Reakcja polimeryzacji trojfosforybonukleotydow na matrycy RNA katalizowana przez rybozym : (a) schemat kompleksu reakcyjnego, (b) analiza produktow reakcji.
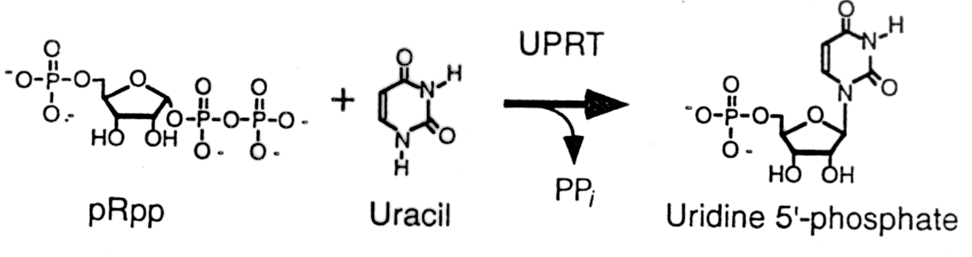
Ryc. 1 - 15. Schemat reakcji syntezy 5' fosforybonukleotydu.
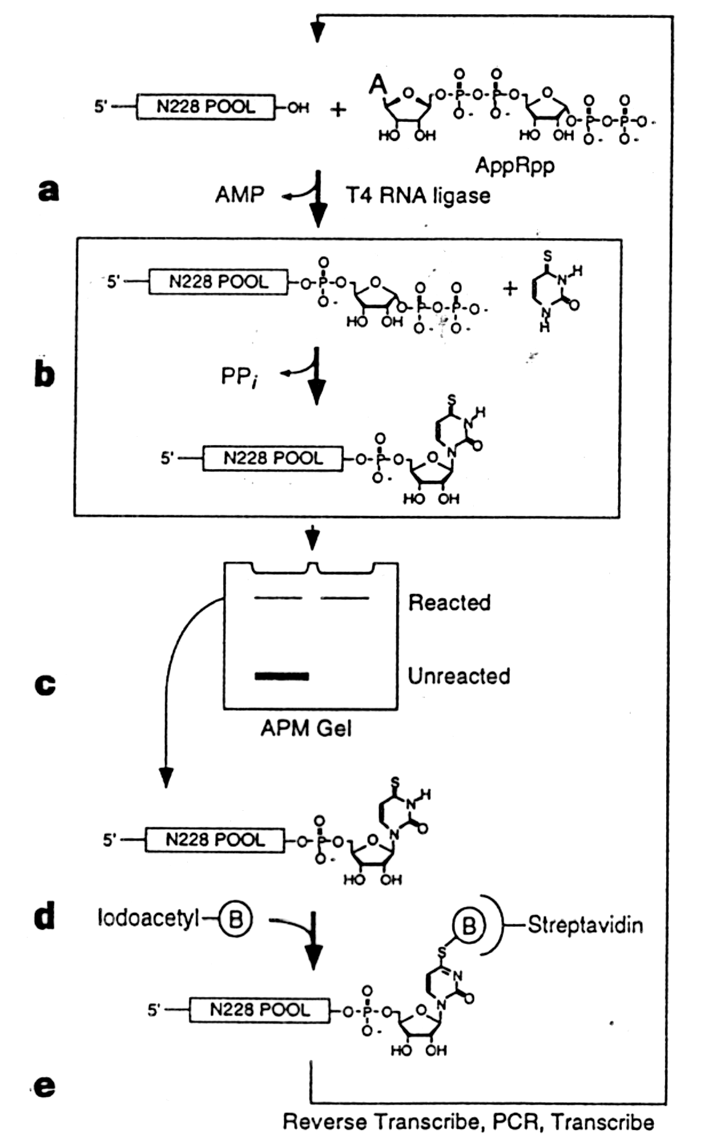
Ryc. 1 - 16. Synteza i selekcja in vitro rybozymu katalizujacego sunteze fosforylowanego rybonukleotydu.
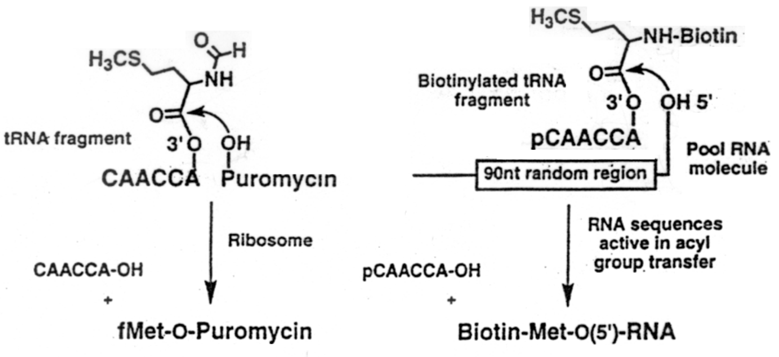
Ryc. 1 - 17. Schemat reakcji hydroksypuromycyny z formylo-metionino-RNA oraz schemat reakcji syntetycznego rybozymu katalizujacego reakcje tworzenia wiazania peptydowago.
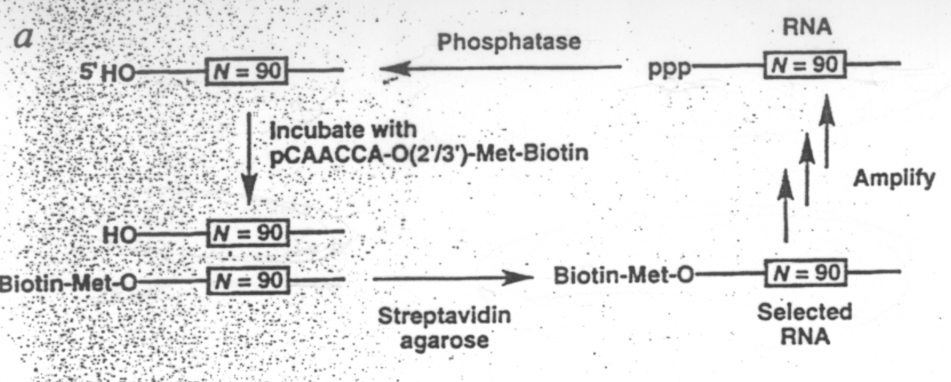
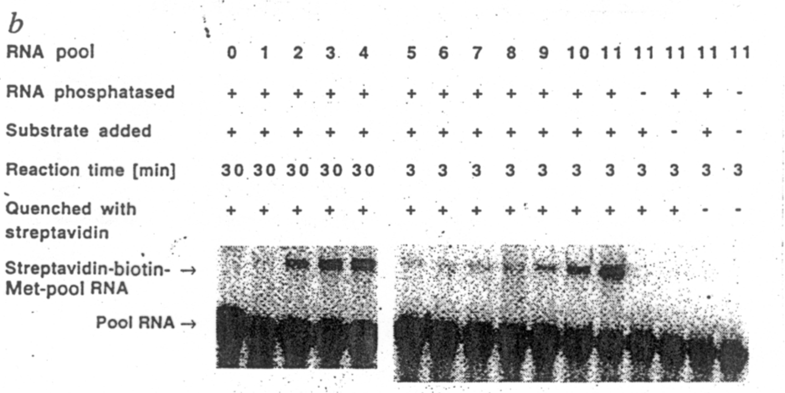
Ryc. 1 - 18. Schemat syntezy i selekcji in vitro rybozymu katalizujacego tworzenie wiazania peptydowego.
Ryc. 1 - 19. Analiza produktow reakcji syntezy wiazania peptydowego katalizowanej
rybozymem otrzymanym w drodze 11 cykli selekcji in vitro.
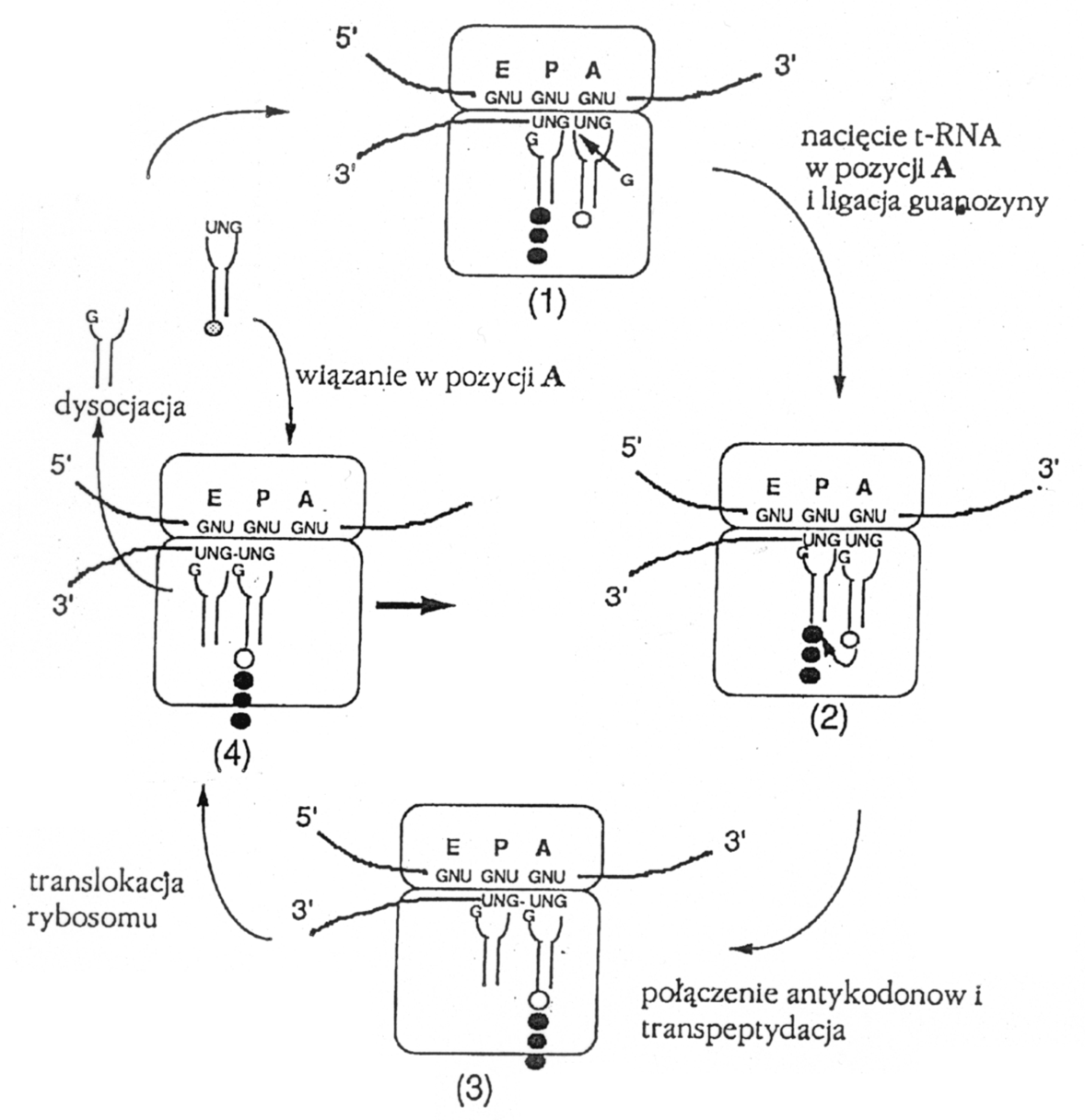
Ryc. 1 - 20. Hipotetyczny mechanizm sprzezenia translacji z replikacja polirybonukleotydu na wczesnym etapie ewolucji.
3
Ryc. 1 - 21. Hipoteza powstania blony komorkowej.
Wyszukiwarka
Podobne podstrony:
01 Ewolucyjne wyselekcjonowanie
01 Rola odchodow dinozaura w sporze ewolucjonizm kreacjonizm (2007)
TD 01
Ubytki,niepr,poch poł(16 01 2008)
Ewolucja marketingu era produkcyjna, sprzedazowa, marketingowa Rynek definicja
01 E CELE PODSTAWYid 3061 ppt
01 Podstawy i technika
01 Pomoc i wsparcie rodziny patologicznej polski system pomocy ofiarom przemocy w rodzinieid 2637 p
zapotrzebowanie ustroju na skladniki odzywcze 12 01 2009 kurs dla pielegniarek (2)
01 Badania neurologicz 1id 2599 ppt
01 AiPP Wstep
ANALIZA 01
01 WPROWADZENIA
Systemy teoretyczne socjologii naturalistycznej – pozytywizm, ewolucjonizm, marksizm, socjologizm pp
01 piątek
więcej podobnych podstron