DSC03040

m VI. Organy rozmnażania Cońiferopsida
samorzutnie. Tylko w części gatunków Taxodiaceae i Tcuaceae znajdowano płytkie komory pyłkowe, nie sięgające głębiej niż 1—2 warstwy komórek powierzchniowych (lit. Singh i Johri 1972, Singh 1978).
Megasporocyt otoczony tkanką gąbczastą rozwija się głęboko w ośrodka w linearną tetradę megaspor. Zasadniczo komórka arche-sporialna powinna się różnicować pod skórką na wierzchołku ośrodka. Niestety rozpoznanie jej wśród innych komórek jest zwykle niemożliwe. Widoczną staje się natomiast jedna z potomnych komórek, kiedy zaczyna się zamieniać w macierzystą komórkę megaspor (megasporocyt); leży ona głęboko w ośrodku, oddzielona od skórki kilkoma warstwami komórek somatycznych. Komórka ta rośnie i przygotowuje się do mejozy. Pochodzenie macierzystej komórki megaspor da się odtworzyć na podstawie układu komórek w tkance ośrodka. W idealnym przypadku na preparacie można znaleźć megasporocyt połączony jednym szeregiem komórek z komórką położoną pod skórką na wierzchołku ośrodka, tj. w miejscu gdzie powinna była być komórka archesporialna. Wnioskujemy stąd, że w rozwijającym się ośrodku komórka archesporialna dzieliła się peryklinalnie raz lub, być może, więcej razy, a następnie w tej samej płaszczyźnie dzieliły się komórki potomne. Powstawał długi szereg komórek, z których ostatnia, najgłębiej zlokalizowana, zmienia pod wpływem jakichś czynników sposób podziału z mi-totycznego na mejotyczny.
W rzeczywistości taki przejrzysty układ komórek w młodym zalążku jest rzadko spotykany, zwykle komórki powstałe z podzielenia się komórki archesporialnej dzielą się w rozmaitych płaszczyznach, co prowadzi do zatarcia jasnego porządku szeregów (rys. 77). Trudno wtedy powiedzieć, gdzie było miejsce komórki archesporialnej. Nie pomoże tutaj oglądanie preparatów z bardzo młodych zalążków, bo często komórka archesporialna niczym zewnętrznie się nie odróżnia od sąsiednich komórek warstwy subepidermalnej. Różnica, jak się zdaje, ujawnia się jedynie w późniejszym rozwoju, bowiem tylko wśród komórek potomnych komórki archesporialnej jedna stanie się mejocytem.
Z komórek wokół macierzystej komórki megaspor rozwija się tkanka odżywcza, nazywana gąbczastą, albo tapetalną. Tkanka ta jest później niszczona, a produkty jej hydrolizy są prawdopodobnie zużywane przez rosnącą megasporę i gametofit. Zanim to nastąpi w megasporocycie odbywa się me-joza (rys. 78).
W końcu mejozy tworzy się zwykle linearna tetrada megaspor, z funkcjonalną megasporą po chalazalnej stronie. Czasami zamiast tetrady rozwija się triada, kiedy w mikropylamej komórce po I podziale mejotycznym nie ma drugiego podziału, a w chalazalnej przebiega on normalnie. Chalazalna me-gaspora triady może dać początek gametofitowi. W zalążku Seguoia setnper-
mrens oprócz linearnego opisano wyjątkowy, izobilaterałny układ megaspor (Looby, Doyle 1942). Również nieczęsto różne gatunki miewają zalążki z dwiema lub większą liczbą macierzystych komórek megaspor. Mogą one przejść mejozę i wydać odpowiednią liczbę tetrad z funkcjonalnymi megaspor ami. Niektóre rozwinięte z nich gametofity dochodzą do pełnej dojrzą-
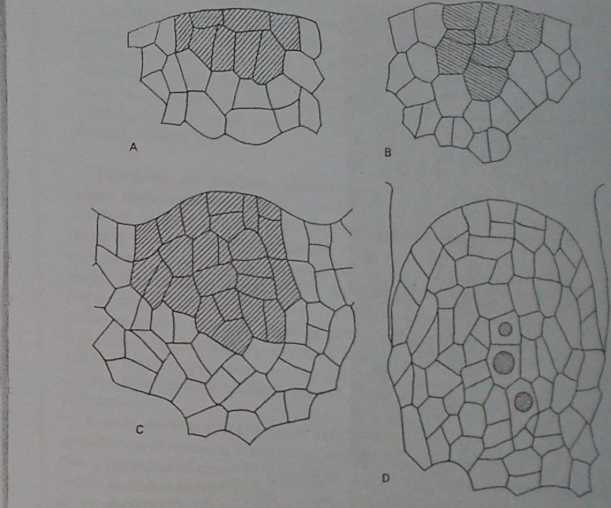
Rys. 77. Inicjacja zalążka sosny (Pimu mago): A — dwie pierwsze warstwy komórek zalążka; B, C — budowa ośrodka, z boków uwypuklają się komórki osłonki; D — ośrodek z macierzystymi ko-mórkami megaspor, z boków zarys brzegu osłonki (Brunkener 1973)
łości, mają archegonia i są zapładniane. Takie nietypowe zalążki z dwoma lub trzema gametofitami w ośrodku są czasem np. u Taxus cmuuknsis, Sarnim, Taxodium i zupełnie wyjątkowo w rodzaju Pinus.
Mejocyt dzieli się asymetrycznie i większość organelli i zapasów wchodzi do megaspory funkcjonalnej. Wyróżnicowany mejocyt w zalążku Chamoecypańs lawsoniana (Cuprcssactae) ma gęstą cytopłazmę z piasty-
Wyszukiwarka
Podobne podstrony:
DSC03026 200 VI. Organy rozmnażania Coniferopsida ścianę komórkową z materiałów zawartych w wytworzo
DSC03028 204 VI. Organy rozmnażania Coniferopsida trzeby komórek linii gen era tywn ej. Przystosowan
DSC03041 230 VI. Organy rozmnażania Coniferopsida darni, niitochondriami i innymi organellami oraz m
DSC03025 VI. Organy rozmnażania W teratologicznej szyszce łuska nasienna może być pędem, a wspierają
DSC03036 #0 VI. Organy rozmnażania Comfaropsida Męskie gamety — komórki plemnikowe, czasem nierówne,
DSC03027 202 VI. Organy rozmnażania Comfempsida ropoleniny pojawiają się, jak sądzimy, najpierw w li
DSC03029 206 VI. Organy rozmnażania ContftropsltU Rys. 68. Rozwój sporangiów iglastych (Comftrae), z
DSC03032 212 VI. Organy, rozmnażania Confcrofistda W cytoplazmie koło nowej plazmolemmy zjawiają się
-organy nadzoru mogą wkraczać w działalność samorządową tylko w przypadkach określonych ustawami: -a
skanowanie0059 196 skie wykazały, że tylko w części przypadków wzbogacenie pracy przyniosło w efekci
herudzinski5 O sposobach rozumienia socjologii wizualnej 27 tylko częścią. Analizowany jest tu świat
pff* & 3 Lotnictwo jest nie tylko częścią sił zbrojnych państwa 1 ważnym czynnikiem gospodę reży
27 - jest on zaspokojony tylko częściowo w przypadku. gdy do momentu podjęcia
VI dotacje od jednostek samorządu terytorialnego w wysokości 3.894.336,47zł, co st
więcej podobnych podstron