DSC03041

230 VI. Organy rozmnażania Coniferopsida
darni, niitochondriami i innymi organellami oraz małe nieliczne wakuole. Większą część miejsca zajmują jednak substancje zapasowe; duże ziania skrobi w amyloplastach i kuleczki tłuszczu wypełniają ściśle oba wierzchołki wydłużonego mejocytu. W okresie I mejotycznej profazy prawie wszystkie mitochondria zbierają się wokół jądra komórkowego, a inne organelle i materiały zapasowe nie zmieniają widocznie miejsca w komórce.
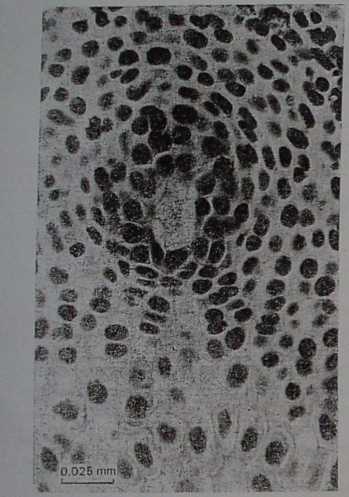
Rys. 78. Zalążek sosny (Pinus sp.), me* gasporocyt w pierwszej profazie mejo* tycznej, kilkanaście dni po zapyleniu (x 800), (fot. A. Hejnowicz)
Mejocyt dzieli się asymetrycznie na większą komórkę chalazalną i mniejszą mikropylarną. Do chalazałncj komórki trafia, jak się zdaje, nieporównanie więcej skrobi i tłuszczów oraz wszystkie, albo prawie wszystkie mitochondria mejocytu. To szczególnie nadaje podziałom znamię zasadniczej nierówności. W powstałej diadzie tylko chalazalna komórka przechodzi II podział męjo-tyczny i daje dwie megaspory ułożone wraz z komórką diady w linearną triadę. Po II podziale mitochondria są widoczne jedynie w chalazałncj megasporze, trudno określić jednak, czy pojedynczych mitochondriów nie
ma w drugiej megasporze, ale jeśli są, to liczba ich musi być nikła. Powstałe komórki triady różnią się potencją rozwoju, co uwidocznia się w chwili, kiedy chalazalna megaspora przekształca się w gametofit, a dwie pozostałe komórki degenerują (Fiordi, Maugini 1977).
Opisane badania w mikroskopie elektronowym potwierdziły i uściśliły dużo wcześniejsze prace zrobione za pomocą mikroskopu świetlnego. Wynikało z nich, że w mejocytach rozmaitych roślin nagozalążkowych (m. in. Taxus, Taxodium} Junipcrus) zaznacza się obszar zagęszczonej cytoplazmy. Od I mejotycznej profazy jest on w chalazalnej części mejocytu, a w niektórych przypadkach po drugim mejotycznym podziale znajduje się w chalazalnej megasporze.
Składniki tej zagęszczonej cytoplazmy nie mogły być dokładnie rozpoznane w mikroskopie świetlnym. Teraz, na podstawie przy toczonych badań w mikroskopie elektronowym możemy sądzić, że główną przyczyną zagęszczenia cytoplazmy jest zebranie się w jednym miejsc?1 dużej j5rytw mitochondriów. Jeżeli to się dzieje w mejocytach, gdzie takie zagęszczenia były widoczne, to u wielu roślin iglastych mitochondria w ciągu megasporo-genezy powinny być wykluczane ze wszystkich komórek pomejotycznych z wyjątkiem chalazalnej megaspory.
Nie wiadomo wcale, jakie czynniki powodują przesunięcia mitochondriów do chalazalnej części mejocytu i utrzymują je w jednym miejscu przez całą mejozę. Jest jednak zrozumiale, że bardzo wielkie skupienie mitochondriów musi wpłynąć na zmianę nasilenia przemiany materii w różnych częściach komórki. Takie zróżnicowanie mejocytu, a później triady lub tetrady można określić jako polaryzację. W komórce (mejocycie) i w układzie komórek są bowiem dwa bieguny różniące się składem organelli i potencjalną zdolnością rozwoju; tylko na biegunie chalazalnym grupują się mitochondria i tylko chalazalna megaspora jest komórką macierzystą gametofitu.
W ścianie mejocytu i tetrady megaspor Chamaecyparis tworzy się gruba warstwa materiału o optycznych cechach kalozy. Związek ten zidentyfikowano cytochemicznie w megasporogenezie innego gatunku Cvpressaam — Biota orientalis (Tarchini 1969). Kaloza występuje jako składnik ściany w mi-kropylamej komórce diady, a prawdopodobnie i w innych stadiach mega-sporogenezy. Z tych dwóch obserwacji trudno wnioskować, czy warstwa ka-lozowa jest odkładana powszechnie w ścianach komórek męjotycznych zalążka roślin iglastych.
Megasporogeneza jest długim procesem, zaczynającym się stopniowo przejściem megasporocytu w I mejotyczną profazę. U Chamtaetypeais z okolic Florencji okres megasporogenczy trwa ze trzy tygodnie marca, oczywiście w innych miejscowościach czas i terminy mogą być inne. Pod koniec men-sporogenezy lub na początku rozwoju gametofitu zalążek najczęściej dojrzewa do zapylenia i wydziela kroplę płynu na mifcropyłe.
Wyszukiwarka
Podobne podstrony:
DSC03026 200 VI. Organy rozmnażania Coniferopsida ścianę komórkową z materiałów zawartych w wytworzo
DSC03028 204 VI. Organy rozmnażania Coniferopsida trzeby komórek linii gen era tywn ej. Przystosowan
DSC03040 m VI. Organy rozmnażania Cońiferopsida samorzutnie. Tylko w części gatunków Taxodiaceae i T
DSC03027 202 VI. Organy rozmnażania Comfempsida ropoleniny pojawiają się, jak sądzimy, najpierw w li
DSC03029 206 VI. Organy rozmnażania ContftropsltU Rys. 68. Rozwój sporangiów iglastych (Comftrae), z
DSC03032 212 VI. Organy, rozmnażania Confcrofistda W cytoplazmie koło nowej plazmolemmy zjawiają się
DSC03025 VI. Organy rozmnażania W teratologicznej szyszce łuska nasienna może być pędem, a wspierają
DSC03036 #0 VI. Organy rozmnażania Comfaropsida Męskie gamety — komórki plemnikowe, czasem nierówne,
230 VI. Fizykalizm zwłaszcza w jego rozważaniach o barwie, gdzie skłonny jest nazwać iluzją to. jako
DSC03006 wegetatywnie■ Charakterystyka genetycznaroślin I rozmnażanych wegetatywnie Potomstwo roślin
230 VI. Fizykaiizm zwłaszcza w jego rozważaniach o barwie, gdzie •skłonny jest nazwać iluzją to. jak
ZASADA OBIEKTYWIZMU ( BEZTRONNOŚCI) ( Art. 4 ) Organy prowadzące postępowanie karne sa obowiązane ba
chemiametale�7 Zadanie 8. Do roztworu siarczanu(VI) chromu(lll) dodano roztwór wodny wodorotlenku so
ORGANY OCHRONY ŚRODOWISKA ■ Marszałek województwa jest organem ochrony środowiska właściwym w
d) Organy, sposób ich powoływania i odwoływania, zakres kompetencji oraz tryb
VI Forum Transportu AglomeracyjnegoWkład prof. Aleksandra Wasiutyńskiego w przekształcenie oraz rozw
Już art. 4 mówi, że organy prowadzące postępowanie kamę są obowiązane badać oraz uwzględniać
Przedsiębiorstwo turystyczne w gospodarce wolnorynkowej G Gołembski (171) 172 VI. Oddziaływanie n
więcej podobnych podstron