DSCN2060 (3)
Powstawanie korzeni bocznych i przybyszowych
Tylko wyjątkowo (u widłaków) korzenie rozgałęziają się widlasto. W tym pr/J, pad ku rozgałęzienie następuje na szczycie korzenia przez utworzenie na miejsce jednego dwu merystemów wierzchołkowych.
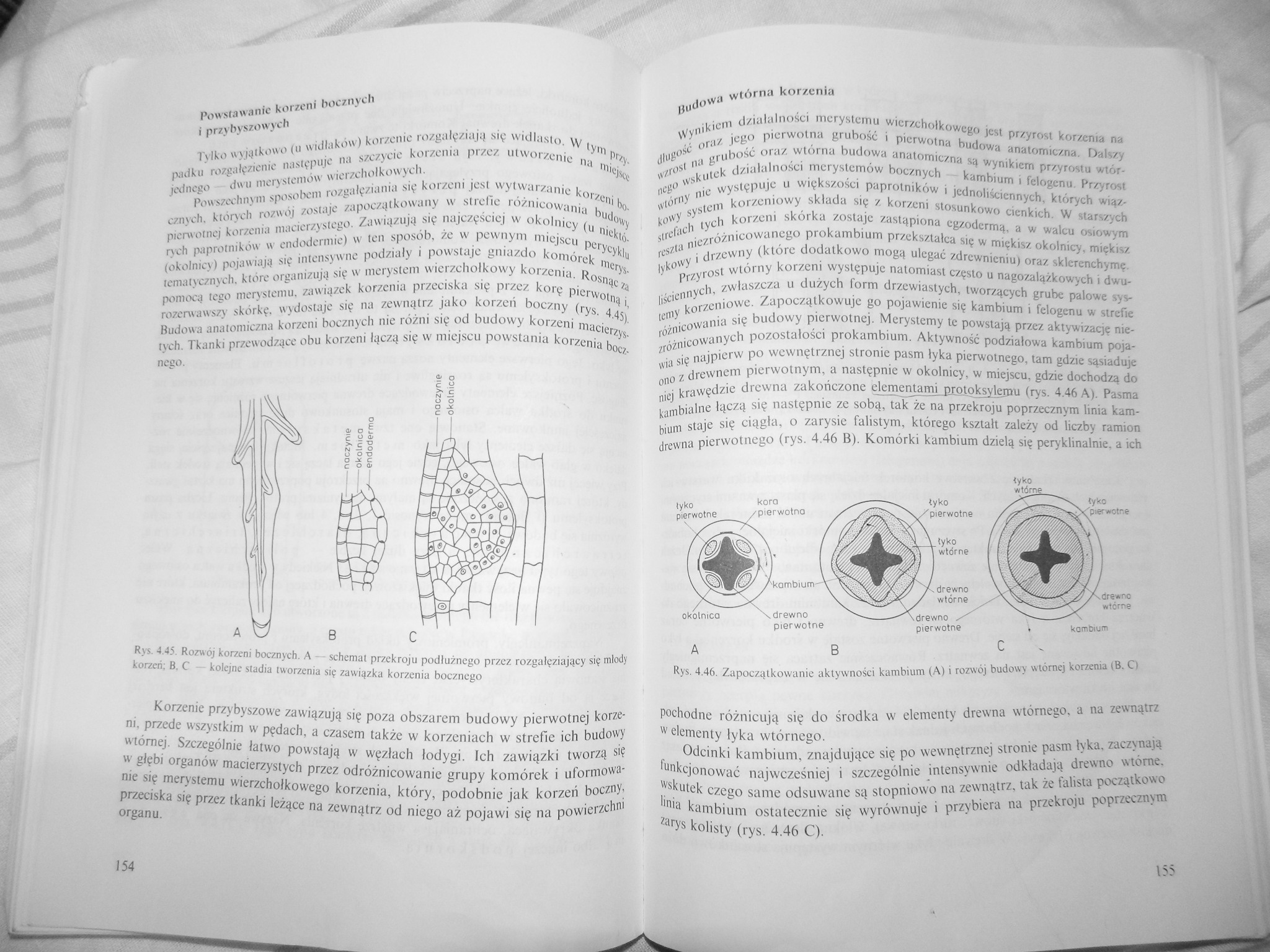
Powszechnym sposobem rozgałęziania się korzeni jest wytwarzanie korzeni bocznych. których rozwój zostaje zapoczątkowany w strefie różnicowania budowy pierwotnej korzenia macierzystego. Zawiązują się najczęściej w okolnicy (u niektórych paprotników w cndoderniie) w ten sposób, że w pewnym miejscu perycyklu (okolnicy) pojawiają się intensywne podziały i powstaje gniazdo komórek merys-tematycznych, które organizują się w merystem wierzchołkowy korzenia. Rosnąc za pomocą tego merystem u. zawiązek korzenia przeciska się przez korę pierwotną i, rozerwawszy skórkę, wydostaje się na zewnątrz jako korzeń boczny (rys. 4.45), Budowa anatomiczna korzeni bocznych nie różni się od budowy korzeni macierzystych. Tkanki przewodzące obu korzeni łączą się w miejscu powstania korzenia bocznego.
Rys. 4.45. Rozwój korzeni bocznych. A — schemat przekroju podłużnego przez rozgałęziający się młody korzeń: B. C - kolejne stadia tworzenia się zawiązka korzenia bocznego
Korzenie przybyszowe zawiązują się poza obszarem budowy pierwotnej korzeni. przede wszystkim w pędach, a czasem także w korzeniach w strefie ich budowy wtórnej. Szczególnie łatwo powstają w węzłach łodygi. Ich zawiązki tworzą się w głębi organów macierzystych przez odróżnicowanie grupy komórek i uformowanie się merystemu wierzchołkowego korzenia, który, podobnie jak korzeń boczny, przeciska się przez tkanki leżące na zewnątrz od niego aż pojawi się na powierzchni organu.
Budowa wtórna korzenia
Wynikiem działalności merystemu wierzchołkowego jest przyrost korzenia na długość oraz jego pierwotna grubość i pierwotna budowa anatomiczna. Dalszy wzrost na grubość oraz wtórna budowa anatomiczna są wynikiem przyrostu wtórnego wskutek działalności merystemów bocznych kambium i felogenu Przyrost wtórny nic występuje u większości paprotników i jcdnoliścicnnych, których wiązkowy system korzeniowy składa się z korzeni stosunkowo cienkich. W starszych strefach tych korzeni skórka zostaje zastąpiona egzodermą, a w walcu osiowym reszta niezróżnicowanego prokambium przekształca się w miękisz okolnicy, miękisz łykowy i drzewny (które dodatkowo mogą ułegać zdrewnieniu) oraz skierenchymę.
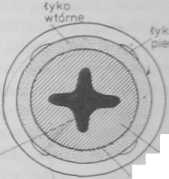
Przyrost wtórny korzeni występuje natomiast często u nagozalążkowych i dwuliściennych, zwłaszcza u dużych form drzewiastych, tworzących grube palowe systemy korzeniowe. Zapoczątkowuje go pojawienie się kambium i felogenu w strefie różnicowania się budowy pierwotnej. Merystemy te powstają przez aktywizację nie-zróżnicowanych pozostałości prokambium. Aktywność podziałowa kambium pojawia się najpierw po wewnętrznej stronie pasm łyka pierwotnego, tam gdzie sąsiaduje ono z drewnem pierwotnym, a następnie w okolnicy, w miejscu, gdzie dochodzą do niej krawędzie drewna zakończone elementami protoksylemu (rys. 4.46 A). Pasma kambialne łączą się następnie ze sobą, tak że na przekroju poprzecznym linia kambium staje się ciągła, o zarysie falistym, którego kształt zależy od liczby ramion drewna pierwotnego (rys. 4.46 B). Komórki kambium dzielą się peryklinalnie. a ich
łyko
koro
pierwotne
\ drewno pierwotne
/pierwotna
kambium

tyko
pierwotne
tyko
wtórne
drewno
wtórne
drewno
B
pierwotne
drewno
wtórne
kambium
A
C
Rys. 4.46. Zapoczątkowanie aktywności kambium (A) i rozwój budowy wtórnej korzenia (B. C)
pochodne różnicują się do środka w elementy drewna wtórnego, a na zewnątrz w elementy łyka wtórnego.
Odcinki kambium, znajdujące się po wewnętrznej stronie pasm łyka, zaczynają funkcjonować najwcześniej i szczególnie intensywnie odkładają drewno wtórne, wskutek czego same odsuwane są stopniowo na zewnątrz, tak że falista początkowo linia kambium ostatecznie się wyrównuje i przybiera na przekroju poprzecznym zarys kolisty (rys. 4.46 C).
Wyszukiwarka
Podobne podstrony:
IMGT39 * pobudzają powstawanie korzeni przybyszowych (auksyny), *
454 SPRAWOZDANIE Z RUCHU REEIGI.INEGO, powinna być wzbronioną, lub dozwoloną tylko wyjątkowo, jeżeli
456 SPRAWOZDANIE Z RUCHU REUIGIJNEGO, tnika wymaga, aby jego żona tylko wyjątkowo do fabryki mogła b
page0384 382 PLATON. czyzną, zwierze organizmem jeszcze niższym. — Gdy w Fedrosie tylko wyjątkowym s
moda kobieca XXw str33 tylko wyjątkowego bogactwa. Dama, o której mowa, nie była wyjątkiem, było po
159 zatem dosięgają jej tylko wyjątkowo silne fale. Na powierzchni plaży woda znajduje się w stanie
P1030023 feremf. Rijkin nej inteligencji i w rezultacie powstał straszny gniot pt. Tylko len jeden r
PICT5424 W klasach I-III ucznia można pozostawić na drugi rok w tej samej klasie tylko w wyjątkowych
77 (211) Badania te, z nielicznymi tylko wyjątkami, wskazują, że uczniowie niepełnosprawni mają gors
110 ARTYKUŁY są w zasadzie książki angielskie, druki obce tylko wyjątkowo. Zarówno wypożyczenia jak
23114 P1030023 feremf. Rijkin nej inteligencji i w rezultacie powstał straszny gniot pt. Tylko len j
0000057 (3) Okołoporodowe uszkodzenie nerwu twarzowego tylko wyjątkowo doprowadza do dłużej trwający
DSC00260 (16) m <2. Sc 3, bu v»aj viwi XI. XII li XI, XII imjdjr no sploty nerwowo powstają; korz
więcej podobnych podstron