
Opisując relacje przestrzenne struktur mózgu stosujemy następujące pojęcia:
brzuszny (ventral) i grzbietowy (dorsal) – oznaczają struktury w odniesieniu
do linii brzuch – plecy u ssaków. Tym samym odnośnie człowieka, „brzuszny”
określa przednią część rdzenia kręgowego od strony brzucha i klatki
piersiowej, zaś w obrębie struktur czaszki te z nich, jakie znajdują się na
przedłużeniu tak wyznaczonej osi, a więc przednich dolnych rejonów mózgu.
„Grzbietowy” natomiast określa tylną część rdzenia kręgowego od strony
pleców i szyi, zaś w obrębie czaszki te struktury, które znajdują się na
przedłużeniu tej osi, a więc tylne i górne rejony półkul mózgu. Pojęcia te są
czasami stosowane do opisu dolnych i górnych jąder danej struktury, jak np.
jądro brzuszne wzgórza.
dziobowy (rostral) i ogonowy (cadual) oznaczają lokalizację albo bliższą
przednich, frontalnych części zwierzęcia, albo bliższą jej tylnych części, na
linii pysk – zadek. W języku polskim nie brzmią idealnie i raczej zastępowane
są innymi.
przedni (anterior) i tylny (posterior) oznaczają lokalizację względem jakiegoś
innego punktu odniesienia, jakiejś innej struktury, wyraźnie bądź domyślnie
określonej. Mogą również opisywać części danej struktury znajdujące się na
linii dziobowy – ogonowy. Przykład: przednia i tylna część zakrętu kory
mózgu; przednia część wzgórza i tylna część wzgórza.
dolny (inferior) i górny (superior) są podobnymi pojęciami jak opisane
powyżej. Przykład: wzgórki górne i dolne; ale odnośnie wzgórza nie mówimy
już o jego górnej, czy dolnej części, stosując w tym przypadku pojęcia
„grzbietowy” i „brzuszny”.
przyśrodkowy (medial) i boczny (lateral) oznaczają struktury na linii
środkowej, uwzględniającej symetryczność mózgu. „Przyśrodkowy” opisuje
lokalizację bliską linii środkowej, znajdującą się na tej części mózgu, jaka nie
jest widoczna z zewnątrz i jaką jedna połówka mózgu styka się z drugą.
„Boczny” odnosi się natomiast do lokalizacji bliższej powierzchni
zewnętrznej.
Opisując budowę wewnętrzną mózgu stosuje się przekroje przez mózg w trzech
płaszczyznach wymienionych poniżej. Oczywiście stosuje się również niestandardowe
przekroje, będące modyfikacją któregoś z poniższych, przeprowadzone pod pewnym kątem
przekrój poziomy (transverse) jest równoległy do podłoża i uwidocznia obie połówki mózgu
przekrój czołowy (coronal) jest prostopadły do podłoża, uwidocznia obie połówki mózgu;
linia przekroju mogłaby przechodzić przez uszy przekrój strzałkowy (sagittal) jest
prostopadły do podłoża, ale w linii prostopadłej do przekroju czołowego, uwidacznia tylko
jedną połówkę mózgu; w linii środkowej przechodzi dokładnie w miejscu, w jakim obie
półkule stykają się.

zarys ogólny
Tkanka nerwowa u naczelnych w zdecydowanej większości wypełnia jamę czaszki i kanał
kręgowy, tworząc zwartą masę określaną mianem ośrodkowego układu nerwowego (o.u.n.; z
ang. układ centralny – central nervous system, CNS). Pozostała tkanka nerwowa, rozsiana
prawie po całym ciele, określana jest mianem obwodowego układu nerwowego (peripheral
nervous system, PNS). Przedmiotem dalszego opisu będzie tylko układ ośrodkowy.
Układ nerwowy jest utworzony z komórek nerwowych, zwanych neuronami, oraz z komórek
glejowych. W dalszej części pominiemy komórki glejowe. Budowę neuronów można w
uproszczeniu sprowadzić do dwóch elementów: (1) dendrytów i ciała komórkowego,
zawierającego jądro komórki i kilka innych jeszcze struktur wewnątrzkomórkowych, oraz (2)
odchodzącego od ciała komórkowego aksonu. Skupiska ciał komórkowych nazywane są
istotą szarą (gray matter), zaś skupiska aksonów – istotą białą (white matter). Fizjologię
przekazywania pobudzeń można w uproszczeniu przedstawić tak, że sygnały dochodzące są
przekazywane na ciało komórkowe, zaś sygnały wychodzące z neuronów są generowane w
miejscu, w jakim akson odchodzi od ciała komórkowego. Tak więc jakkolwiek sygnały
innych neuronów mogłyby wywołać pobudzenie lub hamowanie neuronu, musi mieć to
miejsce jeszcze przed aksonem. Stąd też skupiska istoty szarej traktowane są jako „ośrodki”
przetwarzania informacji, zaś skupiska istoty białej jedynie jako drogi nerwowe. Ośrodki te w
obrębie o.u.n. określamy mianem jąder (nuclei, nucleus), zaś w obrębie układu obwodowego
– zwojów.
Układ ośrodkowy można ogólnie podzielić na następujące struktury:
1. Rdzeń kręgowy (ang. spinal cord).
2. Mózgowie (łac. encephalon):
A. Móżdżek (łac. cerebellum)
B. Pień mózgu (ang. brainstem):
- rdzeń przedłużony, opuszka (łac. medulla oblongata)
- most (łac. pons)
- śródmózgowie (łac. mesoencephalon; ang. midbrain)
C. Mózg (łac. cerebrum):
- międzymózgowie (łac. diencephalon)
- kresomózgowie (łac. telencephalon), czyli półkule mózgu (ang.
cerebral hemispheres).

Rys. Skan MRI w przekroju strzałkowym, ukazujący przyśrodkową powierzchnię jednej z
półkul oraz przekrój innych struktur w linii środkowej: 1 – rdzeń kręgowy; 2 – rdzeń
przedłużony; 3 – most; 4 – śródmózgowie; 5 – wzgórze; 6 – spoidło wielkie; 7 – kora płata
czołowego; 8 – kora płata ciemieniowego; 9 – kora płata potylicznego; 10 – móżdżek.
Kora mózgu, płaszcz istoty szarej okrywający półkule, stanowi najwyższe piętro
przetwarzania informacji. To tutaj ostatecznie docierają sygnały zapoczątkowane w narządach
zmysłu i to stąd wychodzą impulsy nerwowe skierowane następnie do mięśni. Kora mózgu
tworzy najbardziej „ludzkie” struktury umysłu. W opracowywaniu tych informacji pomagają
jej skupiska istoty szarej znajdujące się wewnątrz półkul, oraz wzgórze – największa część
międzymózgowia. Półkule mózgu i centralnie położone wzgórze osadzone są na pniu mózgu,
który pośredniczy w przekazywaniu informacji między narządami zmysłów a korą, oraz
między korą a mięśniami. Samodzielnie też steruje funkcjami wegetatywnymi organizmu,
integrującymi różne systemy fizjologiczne w jedną całość – żywy organizm. Pień mózgu i
rdzeń kręgowy stanowią też wrota o.u.n. względem pozostałego ciała organizmu, z którym
wzajemna komunikacja jest możliwa dzięki połączeniom układu obwodowego. W dalszych
paragrafach skupiono się nie tyle co na opisaniu dokładnej anatomii o.u.n., ile raczej na
zasygnalizowaniu wybranych zagadnień, wspomnieniu o pewnych częściach całego systemu
znajdujących się w określonej okolicy o.u.n. Zagadnienia te ilustrowane są za pomocą
odnośników znajdujących się na samym końcu.
układ komorowy i opony mózgu
Układ komorowy to kilka „pustych” przestrzeni wewnątrz mózgowia, w jakich splot
naczyniówkowy wytwarza płyn mózgowo-rdzeniowy (cerebrospinal fluid, CSF). Każda z
półkul mózgu zawiera komorę boczną (lateral ventricle). Na przekroju strzałkowym komora

boczna ma łukowaty, podłużny kształt odwróconej litery C, od której odchodzą tzw. rogi.
Komory boczne w swych centralnych częściach nieomal stykają się ze sobą, ale rozdziela je
przegroda przeźroczysta (septum pellucidum). Komory boczne łączą się ze sobą małym
otworem pośrodku, gdzie znajduje się pojedyncza komora III (third ventricle), położona
centralnie. Komorę III otacza wzgórze. Płyn mózgowo-rdzeniowy przepływa następnie za
pomocą wodociągu mózgu (aqueduct of Sylvius) do komory IV (fourth ventricle), znajdującej
się grzbietowo na pniu mózgu i graniczącej z móżdżkiem. Należy podkreślić, że komora III i
IV są strukturami pojedynczymi, leżącymi w linii centralnej. Płyn mózgowo-rdzeniowy
wydostaje się z komory IV do przestrzeni podpajęczynówkowej (subarachnoid space),
zawartej między trzema oponami mózgu (meninges). Są one tkankami otaczającymi
mózgowie i rdzeń kręgowy, zaś płyn dzięki temu może okrążać cały ośrodkowy układ
nerwowy. Opony oddzielają mózg od kości czaszki, oraz rdzeń kręgowy od kości kręgosłupa.
Choroby degeneracyjne, w wyniku których postępuje masowa śmierć neuronów, prowadzą w
naturalny sposób do zmniejszania się masy mózgowia i powiększania się komór mózgu (co
uwidacznia się w neuroobrazowaniu). W dawnych czasach niektórzy filozofowie traktowali
komory mózgu jako siedlisko duszy.
rdzeń kręgowy
Rdzeń kręgowy jest tworem podłużnym i prostopadłym do podłoża, stosunkowo cienkim o
średnicy ok. 1 cm. Rdzeń można podzielić na poszczególne odcinki ulokowane jeden na
drugim. Każdy taki odcinek wysyła aksony do odpowiadającej mu części ciała unerwiając ją
ruchowo, jak i otrzymuje z tej części aksony rozlokowanych tam komórek czuciowych.
Odcinki te noszą nazwę segmentów szyjnych (cervical; C1-C8), piersiowych (thoracic; Th1-
Th12), lędźwiowych (lumbar; L1-L5) i krzyżowych (sacral; S1-S5). Jako ostatni występuje
jeszcze jeden odcinek guziczny (coccygeal). Podobnie określa się poszczególne segmenty
kręgosłupa, jednakże fizycznie nie odpowiadają sobie, ponieważ rdzeń jest krótszy i wypełnia
kanał kręgowy jedynie w części. Rdzeń kończy się mniej więcej na wysokości pierwszych
kręgów lędźwiowych kręgosłupa. Dalej biegną już włókna nerwowe nazywane ogonem
końskim (cadua equina), związane z odcinkami lędźwiowymi, krzyżowymi i guzicznym
rdzenia, które opuszczają kanał kręgowy we właściwych sobie miejscach. Podczas gdy w
całym o.u.n. dominuje jednolita organizacja istoty szarej i białej – istota szara jest na
zewnątrz, zaś biała wypełnia wnętrze, rdzeń kręgowy jest zorganizowany na odwrót. Jego
boczne części tworzone są przez drogi nerwowe (aksony) przesyłające informacje między
mózgowiem a rdzeniem kręgowym w obu kierunkach. Otaczają one jądra istoty szarej
zapewniające podstawowe łuki odruchowe i stacje przekaźnikowe na drodze przekazywania
informacji. Istota szara rdzenia w obrębie każdego segmentu tworzy wystające ze środka dwie
klasy rogów. Rogi brzuszne (ventral horn) zawierają przede wszystkim jądra motoryczne i
wysyłają na obwód nerwy ruchowe. Do rogów grzbietowych (ang. dorsal horn) natomiast
wnikają nerwy czuciowe. Uszkodzenie rdzenia kręgowego przede wszystkim przerywa drogi
nerwowe, czego konsekwencją jest niemożność sterowania mięśniami przez korę mózgu lub/i
niemożność przez nią odbierania informacji czuciowej (paraliż – plegia, niedowład – paresis,

niedoczulica). Uszkodzenie tych dróg gdziekolwiek one przebiegają (w mózgowiu, na
obwodzie) również prowadzi do takich konsekwencji.
móżdżek
Móżdżek jest złożony z dwóch półkul móżdżku (cerebellum hemispheres) oraz położonego
centralnie robaka (vermis). Od strony zewnętrznej móżdżek jest pokryty trzema warstwami
istoty szarej, tworzącymi niezwykle pofałdowaną korę móżdżku. Najwięcej uwagi
przyciągają neurony Purkynego (lub Purkyniego), mające – co bardzo charakterystyczne –
niezwykle rozbudowane i gęste drzewka dendrytyczne. Najwyższa warstwa kory móżdżku
utworzona jest po przez dendryty tych komórek, zaś ich ciała komórkowe ulokowane są w
drugiej warstwie. Informacje dochodzące zmierzają właśnie do dendrytów komórek
Purkynego. Wewnątrz istoty białej półkul móżdżku znajdują się podkorowe skupiska istoty
szarej – jądra móżdżku (cerebellar nuclei): jądro zębate (denate), jądro wsunięte (interpositus)
– czopowate (emboliform) i kulkowate (globose), oraz jądro wierzchu (fastigal). Jądra
móżdżku m.in. pośredniczą w przekazywaniu informacji z komórek Purkynego poza
móżdżek. Móżdżek jest intensywnie połączony ze rdzeniem kręgowym, korą ruchową półkul
mózgu i wzgórzem. Jego funkcje można ogólnie określić jako związane z kontrolą czynności
mięśni. Konsekwencją uszkodzenia móżdżku jest ataksja móżdżkowa, polegająca na utracie
płynności ruchów, które stają się niezborne, co szczególnie uwidacznia się przy ruchach
zamierzonych kończyn, jak również dyzartria móżdżkowa, objawiająca się mową
skandowaną.
pień mózgu
Ogólnie
Jeżeli spojrzeć na układ nerwowy od spodu, pień mózgu jest jak gdyby kontynuacją rdzenia
kręgowego, ale im wyżej, tym bardziej wzrasta złożoność budowy i pojawia się coraz więcej
nowych elementów.
Kontynuacją „rdzeniowych” funkcji zajmują się jądra nerwów czaszkowych (cranial nerves).
12 par nerwów czaszkowych unerwia ruchowo i czuciowo m.in. mięśnie twarzy, krtani,
języka i gałki ocznej. Uszkodzenie jąder w pniu bądź nerwów czaszkowych na obwodzie
powoduje niedowład tych mięśni lub/i niedoczulicę. Pień mózgu pośredniczy w odbiorze i
przekazywaniu do kory informacji zmysłu słuchu i smaku, znajdują się tu również stacje

przekaźnikowe na drodze przesyłania informacji czuciowych (dotyk, temperatura, ból,
wibracje) z rdzenia. W obrębie właściwie całego pnia mózgu rozsiany jest twór siatkowaty
(reticular formation), kontrolujący m.in. aktywność mięśniową i wywierający nieswoisty
wpływ pobudzający na mózgowie. Dzięki temu kontroluje on stan snu i czuwania. Do tworu
siatkowatego należą m.in. jądra miejsca sinawego (o czym będzie jeszcze wspomniane), czy
jądra szwu.
Rdzeń przedłużony
Rdzeń przedłużony (medulla oblongata) jest związany z nerwami czaszkowymi IX
(językowo-gardłowy), X (błędny), XI (dodatkowy) i XII (podjęzykowy). Odpowiadają one
m.in. za aktywność gruczołów ślinowych, odruchy połykania, ruchy języka i krtani, unerwiają
kubki smakowe i struktury ucha. Biorą tym samym udział w mówieniu, przełykaniu i
oddychaniu. Za pomocą tych nerwów przenoszone są również istotne dla życia informacje
układu autonomicznego (wegetatywnego). Np. jądro dwuznaczne (nucleus ambiguus), które
jest motorycznym jądrem dla nerwów IX, X i XI, wysyła również włókna autonomiczne do
zwojów sercowych. Można ogólnie powiedzieć, że rdzeń przedłużony zawiera ośrodki
wegetatywne (vegetative centers) odpowiedzialne za sterowanie akcją serca, oddychaniem,
jak również motoryką żołądkowo-jelitową. Uszkodzenie tych ośrodków, określane mianem
śmierci pnia mózgu, oznacza definitywnie zgon zgon (nawet jeśli możliwe jest sztuczne
podtrzymywanie krążenia). Warto również wspomnieć, że w obrębie rdzenia przedłużonego
znajduje się skrzyżowanie drogi korowo-rdzeniowej. Droga ta jest zapoczątkowana w korze
motorycznej półkul mózgu i przesyła pobudzenia mięśni do rdzenia kręgowego, skąd
informacje są przekazywane już bezpośrednio mięśniom po przez układ obwodowy. Włókna
te na poziomie rdzenia przedłużonego tworzą charakterystyczne struktury nazywane
piramidami (pyramids). Skrzyżowanie piramid (decussation of pyramids) to miejsce, w jakim
aksony zapoczątkowane w danej półkuli mózgu przechodzą na przeciwną stronę układu
nerwowego, tak, że prawa półkula steruje lewą stroną ciała, zaś lewa półkula zarządza prawą
stroną ciała.
Most
Most (pons) jest związany z nerwami czaszkowymi V (trójdzielny), VI (odwodzący), VII
(twarzowy), oraz VIII (przedsionkowo-ślimakowy). Nerwy te unerwiają gruczoły ślinowe i
łzowe, gałki oczne, mięśnie twarzy, zbierają informacje słuchowe, smakowe, czuciowe z
obrębu twarzy, biorą udział w utrzymaniu równowagi. Most odbiera informacje słuchowe
wygenerowane przez ślimaka. Do mostowych ośrodków i tworów istoty białej, związanych ze
zmysłem słuchu zaliczamy jądra ślimakowe grzbietowe (dorsal cochlear nucleus) i brzuszne
(ventral cochlear nucleus), ciało czworoboczne (trapezoid body), jądro górne oliwki (inferior
olivary nucleus), oraz wstęgę boczną (lateral lemniscus). Ruchy oczu, które są sterowane
zarówno przez most, jak i śródmózgowie, na poziomie mostu są zarządzane przez jądro nerwu
VI odwodzącego (abducent nucleus). Jest ono nazywane również „mostowym ośrodkiem
skojarzonego spojrzenia w bok”, ze względu na funkcję oznaczoną w tej nazwie. Jądro to jest
odbiorcą aksonów korowego ośrodka skojarzonego spojrzenia w bok w płacie czołowym
(frontal eye field, FEF). Uszkodzenia mostu mogą więc prowadzić do zaburzeń związanych z

ruchami oczu, takich jak np. oczopląs, czyli mimowolne i rytmiczne drgania gałek ocznych w
kierunku poziomym, bądź pionowym (ale nie w tym przypadku), ale również do głuchoty,
czy porażenia twarzy. W wyniku znacznego uszkodzenia mostu obejmującego drogi
przechodzące przez most, łączące korę mózgu z rdzeniem kręgowym, jak i nerwy czaszkowe,
rozwija się zespół śpiączki rzekomej, nazywany także zespołem zamknięcia (locked in).
Pacjent cierpi na porażenie czterokończynowe i może się komunikować z otoczeniem jedynie
po przez mruganie i pionowe ruchy oczu. Warto wspomnieć o jeszcze jednej strukturze
mostu. Neurony miejsca sinawego (locus coeruleus) zawierają melaninę (barwnik) i rozsyłają
swoje aksony do pozostałych części mózgowia. Zakończenia synaptyczne tych aksonów jako
neuroprzekaźnika używają noradrenaliny.
Śródmózgowie
Śródmózgowie (midbrain) jest związane z nerwami czaszkowymi III (okoruchowy) i IV
(bloczkowy). Unerwiają one mięśnie powieki, gałki ocznej i źrenicy, biorą udział w
akomodacji. Uszkodzenie tych nerwów i ich jąder może doprowadzić do trudności z
akomodacją, usztywnieniu się źrenicy (która może być stale rozszerzona), diplopii –
podwójnego widzenia przy spoglądaniu na stronę uszkodzenia, czy do pionowego zeza
rozbieżnego. Wśród innych struktur śródmózgowia należy wspomnieć jeszcze o wzgórkach
czworaczych i istocie czarnej. Wzgórki czworacze dolne (inferior colliculus) są stacją
przekaźnikową na drodze przesyłania informacji słuchowych. Ośrodki słuchowe mostu, które
odbierają informację ze ślimaka, przekazują ją do wzgórków dolnych. Aksony wzgórków
dolnych przekazują tę informację do wzgórza w międzymózgowiu, skąd trafia ona już do
pierwszorzędowej kory słuchowej w płacie skroniowym półkuli mózgu. Wzgórki czworacze
górne (superior colliculus) otrzymują natomiast informacje wzrokowe z siatkówki. Nie są
jednak stacją przekaźnikową na drodze informacji wzrokowych miedzy receptorami a korą
mózgu, jak miało to miejsce w przypadku wzgórków dolnych i zmysłu słuchu. W tym
przypadku siatkówka przesyła pobudzenia do wzgórza międzymózgowia a stamtąd
informacje docierają do pierwszorzędowej kory wzrokowej mózgu. Wzgórki górne otrzymują
informacje wzrokowe niejako w sposób niezależny od drogi siatkówkowo-korowej.
Uczestniczą w identyfikacji ruchu w polu wzrokowym i w jego śledzeniu. Ponieważ wzgórki
górne i dolne są wzajemnie połączone, śródmózgowie zapewnia łuki odruchowe dla prostych
odruchów słuchowo-wzrokowych (czy też motoryczno-słuchowo-wzrokowych), jakie mogą
zachodzić bez udziału kory półkul mózgu. Istota czarna (substantia nigra) zawiera w części
zbitej grupę neuronów z melaniną (barwnik). Przesyłają one aksony do podkorowych skupisk
istoty szarej w półkulach mózgu (czyli do jąder podstawy) i używają jako neuroprzekaźnika
dopaminy. Uszkodzenie tej drogi i redukcję neuronów dopaminergicznych istoty czarnej
obserwuje się w chorobie Parkinsona.

mózg
Międzymózgowie: podwzgórze
Podwzgórze (hypothalamus) anatomicznie jest częścią międzymózgowia. Jest to niewielkie
skupisko licznych jąder, ulokowane do przodu od śródmózgowia, w bliskim sąsiedztwie
skrzyżowania nerwów wzrokowych (optic chiasm) oraz przysadki mózgowej (hypophysis;
pituitary gland), organu wydzielania wewnętrznego. Od góry graniczy z dnem komory III.
Funkcjonalnie podwzgórze pełni rolę w administrowaniu homeostazą i układem
autonomicznym (wegetatywnym), mając wpływ na regulację krążenia, temperatury ciała,
pragnienia, uczucia głodu, popędu seksualnego itd. Niektóre z jąder mają też wpływ na
produkowanie i uwalnianie do krwi różnych hormonów, głównie po przez interakcje z
przysadką mózgową. Większość jąder podwzgórza nosi nazwy o charakterze anatomicznym,
np. jądro grzbietowo-przyśrodkowe, jądro tylne, jądro nadskrzyżowaniowe. Prawdopodobnie
najbardziej znanymi jądrami podwzgórza są ciała suteczkowate, oraz jądra pola
przedwzrokowego. Ciało suteczkowate (mammilliary body) jest związane z procesem
zapamiętywania nowych informacji, choć nie wiadomo jaką dokładnie rolę pełni w tym
procesie. Otrzymuje informację z hipokampa, a jego degenerację obserwuje się w
poalkoholowym zespole amnestycznym. Natomiast pole przedwzrokowe (preoptic area)
zawiera cztery jądra INAH (interstitial nuclei of the anterior hypothalamus), z których
najwięcej emocji budzi INAH3, wykazujące zróżnicowanie anatomiczne między płciami oraz
między osobami homo- i heteroseksualnymi. Jest odpowiednikiem jądra SDN – POA
(sexually dimorphic nucleus of the preoptic area – płciowo zróżnicowane jądro pola
przedwzrokowego) u szczura, który to gatunek jest najczęściej przedmiotem badań.
Międzymózgowie: wzgórze
Wzgórze (thalamus) jest inną częścią międzymózgowia. Jest to zwarta struktura jąder
otaczająca komorę III i leżąca powyżej podwzgórza. Jest też znacznie większa niż
podwzgórze. Nazwy jąder wzgórza także mają uwarunkowanie anatomiczne, np. jądro
przednie, brzuszno-boczne, brzuszno-tylne, czy śródblaszkowe (blaszki to skupiska istoty
białej we wzgórzu). Wyjątek stanowi poduszka (ang. pulvinar), największe jądro wzgórza,
oraz ciała kolankowate. Te ostatnie, z punktu widzenia anatomicznego, przynależą do
zawzgórza, jednak zwykło się traktować je łącznie ze wzgórzem, aby nie mnożyć bytów
ponad miarę. Ciała kolankowate boczne (lateral geniculate nucleus, LGN) są związane ze
wzrokiem, otrzymują informację z siatkówki i przekazują ją następnie do kory półkul mózgu.
Ciała kolankowate przyśrodkowe (medial geniculate nucleus, MGN) są związane ze słuchem,
otrzymują informacje słuchowe z pnia mózgu, a dalej przekazują już do kory mózgu.
Pozostałe jądra wzgórza są związane z innymi zmysłami, za wyjątkiem jąder
śródblaszkowych oraz poduszki, które to jądro pełni funkcje integracyjne dla informacji
różnych zmysłów. Wzgórze posiada wzajemne połączenia z korą półkul mózgu. Właściwie
każda okolica korowa ma swoje jądro we wzgórzu.

Kresomózgowie
Kresomózgowie, czyli półkule mózgu, budzą najwięcej emocji i zainteresowania. Dlatego też
to im przeznaczymy najwięcej miejsca (a szczególnie zaś korze niejednorodnej i nowej).
Istota szara półkul:
1. kora mózgu (cortex):
a. sześciowarstwowa kora nowa, jednorodna (neocortex; isocortex)
b. trójwarstwowa kora niejednorodna (allocortex: archicortex,
paleocortex)
c. sześciowarstwowa kora przejściowa (mesocortex)
2. struktury podkorowe, tzw. jądra podstawy (basal ganglia):
a. przedmurze (claustrum)
b. ciało migdałowate (amygdala)
c. gałka blada (globus pallidus)
d. prążkowie grzbietowe (striatum)
- skorupa (putamen)
- jądro ogoniaste (caudate nucleus)
e. prążkowie brzuszne – jądro półleżące (nucleus accumbens)
f. jądro podstawne Meynerta.
Istota biała półkul tworzy:
1. Włókna wewnątrzpółkulowe, łączące ze sobą różne okolice tej samej półkuli.
Składają się na nie krótkie, łukowate włókna łączące sąsiadujące zakręty, oraz
dłuższe włókna nazywane pęczkami, łączące różne okolice jednej półkuli.
2. Włókna międzypółkulowe, spoidłowe, które łącząc obie półkule tworzą tzw.
spoidła w linii środkowej:
o
spoidło wielkie, ciało modzelowate (corpus callosum)
o
spoidło przednie (anterior commissure)
o
spoidło hipokampa (fornix).
3. Włókna rzutowe, nazywane także projekcyjnymi, łączące półkulę mózgu z
innymi (niższymi) obszarami układu nerwowego, które zbiegając się razem
tworzą tzw. wieniec promienisty (corona radiata), zaś przechodząc między
jądrami podstawy i korą mózgową tworzą "torebki" (capsule).
Kora otaczająca półkule to płaszcz ciał komórkowych neuronów. Kora mózgu jest
pofałdowana, tworzy uwypuklenia – zakręty (gyri, gyrus), oraz szczeliny – bruzdy (fissure,
sulci, sulcus). Stosunkowo duża część kory zawija się w głąb bruzd. Zasadniczo w każdym
zdrowym mózgu zakręty i bruzdy układają się w typowy sposób, choć z pewną dozą
indywidualności. Każda półkula jest dzielona na cztery płaty (lobes): czołowy (frontal),
ciemieniowy (parietal), potyliczny (occipital) i skroniowy (temporal). Wyróżnia się jeszcze

wyspę (insula), lub płat wyspowy, który jest sporym fragmentem pofałdowanej kory ukrytej
w bruździe bocznej, niejako wepchniętym do środka półkuli. Czasami funkcjonuje jeszcze
tradycyjny termin płat limbiczny (limbic), określający pewne struktury ulokowane na
powierzchni przyśrodkowej (jak hipokamp i zakręt obręczy).
Rys. Szkic powierzchni bocznej jednej z półkul. Wyższy rysunek ukazuje podział na płaty,
dolny rysunek przedstawia zakręty: 1 – górny zakręt czołowy (superior frontal gyrus); 2 –
środkowy zakręt czołowy (middle frontal gyrus); 3 – dolny zakręt czołowy (inferior frontal
gyrus); 4 – kora oczodołowa (orbital cortex); 5 – zakręt przedśrodkowy (precentral gyrus); 6 –
zakręt zaśrodkowy (postcentral gyrus); 7 – zakręt nadbrzeżny (supramarginal gyrus); 8 –
zakręt kątowy (angular gyrus); 9 – płacik ciemieniowy dolny (tj. zakręt nabrzeżny + kątowy
inferior parietal lobule); 10 – płacik ciemieniowy górny (superior parietal lobule); 11 –
zakręty potyliczne boczne (lateral occipital gyri); 12 – górny zakręt skroniowy (superior
temporal gyrus); 13 – środkowy zakręt skroniowy (middle temporal gyrus); 14 – dolny zakręt
skroniowy (inferior temporal gyrus).
Kora mózgu jest pofałdowana, a każdy fałd (zakręt) i bruzda posiada swoją nazwę. Spośród
najważniejszych bruzd na powierzchni bocznej należy wspomnieć o bruździe środkowej
(central sulcus), przebiegającej prostopadle do podłoża i rozdzielającej płaty czołowy i
ciemieniowy, oraz o bruździe bocznej (lateral sulcus), biegnącej bardziej równolegle do
podłoża i oddzielającej płat skroniowy od płatów czołowego i ciemieniowego. Dodatkowo na
powierzchni przyśrodkowej, co już nie jest ukazane na powyższej ilustracji, w obrębie płata
ciemieniowego przebiega bruzda ostrogowa (calcarine sulcus). Znajduje się ona na linii
biegun płata potylicznego – centrum półkuli. Kora znajdująca się nad bruzdą ostrogową
określana jest mianem klinka (cuneus), zaś kora pod tą bruzdą – mianem zakrętu potyliczno-
skroniowego przyśrodkowego (medial occipitotemporal gyrus) lub zakrętu językowatego
(lingual gyrus). Na powierzchni przyśrodkowej znajduje się również zakręt obręczy (cingulate
gyrus), przebiegający łukowato nad spoidłem wielkim w obrębie płatów czołowego i
ciemieniowego. Zakręty widoczne na powierzchni bocznej półkul są ukazane na poniższej
ilustracji.

Rys. Szkic powierzchni bocznej jednej z półkul: 1 – górny zakręt czołowy (superior frontal
gyrus); 2 – środkowy zakręt czołowy (middle frontal gyrus); 3 – dolny zakręt czołowy
(inferior frontal gyrus); 4 – kora oczodołowa (orbital cortex); 5 – zakręt przedśrodkowy
(precentral gyrus); 6 – zakręt zaśrodkowy (postcentral gyrus); 7 – zakręt nadbrzeżny
(supramarginal gyrus); 8 – zakręt kątowy (angular gyrus); 9 – płacik ciemieniowy dolny (tj.
zakręt nabrzeżny + kątowy inferior parietal lobule); 10 – płacik ciemieniowy górny (superior
parietal lobule); 11 – zakręty potyliczne boczne (lateral occipital gyri); 12 – górny zakręt
skroniowy (superior temporal gyrus); 13 – środkowy zakręt skroniowy (middle temporal
gyrus); 14 – dolny zakręt skroniowy (inferior temporal gyrus).
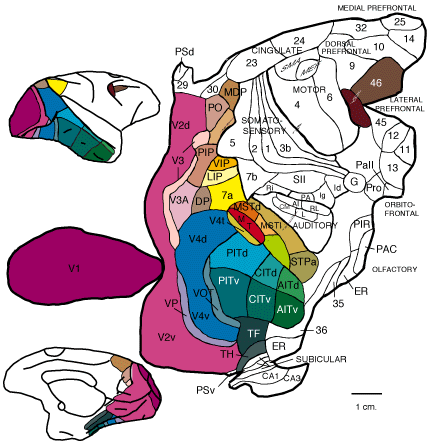
Oprócz terminów stricte anatomicznych, dotyczących nazw poszczególnych bruzd i zakrętów,
stosuje się także i inne określenia. Kora mózgu może być dzielona na okolice, które
wyróżniają się szczegółami budowy istoty szarej od obszarów sąsiednich. Budowę struktur
tworzonych przez istotę szarą określa się mianem cytoarchitektoniki. W przypadku mózgu
człowieka najbardziej popularną mapą cytoarchitektoniczną jest podział niemieckiego
anatoma Korbiniana Brodmanna (1868-1918), opublikowany poraz pierwszy w 1909 roku. W
tym przypadku okolice kory mózgu nie mają nazw, ale numery (Brodmann area, BA).
Czytelnik może obejrzeć sobie tę mapę w Internecie, np. tutaj 1 | 2 | 3 | 4 | 5 (makak).
Innym sposobem podziału okolic korowych jest wyróżnianie ośrodków mających odrębne
cechy fizjologiczne. Tutaj nazewnictwo jest często wymieszane, pochodzące od klasyfikacji
różnych autorów i nawet pojedynczych prac opisujących wytyczanie granic poszczególnych
okolic. Zastosowanie ma najczęściej terminologia anatomiczna, np. MT – okolica środkowa
skroniowa (middle temporal) – określa pewien obszar kory związany z przetwarzaniem
informacji wzrokowych. Obecnie można spotkać również płaskie mapy kory, ukazujące
jednocześnie powierzchnie boczą i przyśrodkową jednej z półkul. Zazwyczaj właśnie na
takich mapach przedstawia się podział na okolice czynnościowe. Na przełomie lat 80. i 90.
David C. Van Essen, w oparciu o serię wymierzonych histologicznych skrawków szesnastu
poziomych przekrojów mózgu małpy utworzył tego typu mapę bodajże po raz pierwszy.
Celem tej żmudnej procedury było dokładne przedstawienie różnych okolic wzrokowych,
jakich granice wyznaczono wówczas w licznych eksperymentach fizjologicznych i
anatomicznych. Trudno jest je przedstawić na standardowej, rzeczywistej ilustracji chociażby
z tego względu, że większość z nich jest ukryta wewnątrz różnych bruzd. Tymczasem płaska
mapa pozwala „wyprasować” powierzchnię półkuli z bruzd i zakrętów i uzmysłowić jak wiele
kory jest ukrytej w bruzdach. Obecnie mapowanie poszczególnych okolic wzrokowych
człowieka i małp odbywa się za pomocą fMRI, zaś płaskie mapy tworzy się z użyciem metod
obliczeniowych i stosownego oprogramowania. Mimo odmiennego sposobu ukazania
powierzchni kory, płaskie mapy nadal odwołują się do pól Brodmanna. Mapy te czytelnik
może obejrzeć w Internecie, np. tutaj: 1 | 2 | 3 | 4 | 5 (makak).

Choćby pobieżne omówienie funkcji półkul mózgu wymaga odrębnego artykułu i tak też
uczyniliśmy. Tutaj jedynie zostanie zarysowane to zagadnienie w stopniu, jaki być może
pozwoli lepiej poznać budowę półkul mózgu. Generalnie – kora półkul mózgu stanowi
najwyższe piętro przetwarzania informacji, jest siedliskiem mechanizmów odpowiedzialnych
za te funkcje, które w największym stopniu uznajemy jako umysłowe, mentalne, psychiczne.
To, że możemy się swobodnie poruszać, widzieć, słyszeć, mówić i pamiętać, zawdzięczamy
skomplikowanym obwodom korowym, których uszkodzenie może pozbawić nas subtelnych
składników tych procesów, a nawet i w ogóle całych funkcji. Chcielibyśmy zwrócić uwagę
czytelnika na kilka wybranych, następujących struktur kory mózgu:
Pierwszorzędowa kora wzrokowa V1 (primary visual cortex) – kora płata potylicznego
wewnątrz i wokół bruzdy ostrogowej (calcarine sulcus), przebiegającej na powierzchni
przyśrodkowej. Pokrywa się z 17 polem Brodmanna. Jest to okolica odbierająca informacje
wzrokowe ze wzgórza.
Pierwszorzędowa kora słuchowa A1 (primary auditory cortex) – zakręty poprzeczne
Heschla, ulokowane w środkowej części górnego zakrętu skroniowego, przebiegające
prostopadle do jego linii (stąd nazwa) w jego górnej powierzchni i wnikające w głąb bruzdy
bocznej. Odpowiadają polom 41 i 42 Brodmanna. Jest to okolica odbierająca informacje
słuchowe ze wzgórza.
Pierwszorzędowa kora czuciowa S1 (primary somatosensory cortex) – zakręt zaśrodkowy,
przebiegający równolegle do bruzdy środkowej (central sulcus) i prostopadle do podłoża w
płacie ciemieniowym. Pokrywa się z polami 1, 2 i 3 Brodmanna. Jest to okolica odbierająca
informacje somatosensoryczne ze wzgórza.
Pierwszorzędowa kora ruchowa M1 (primary motor cortex) – zakręt przedśrodkowy,
równoległy do bruzdy środkowej (central sulcus) i biegnący prostopadle do podłoża w płacie
czołowym. Pokrywa się z polem 4 Brodmanna. Jest to okolica wysyłająca aksony drogą
korowo-wzgórzową.
Ośrodek ruchowy mowy (Broca) – tylna część dolnego zakrętu czołowego w lewej półkuli,
pokrywająca się z polami 45 i 44 Brodmanna. Jest to okolica związana z nadawaniem mowy,
która pod tym kątem steruje korą ruchową dla mięśni aparatu artykulacyjnego.
Ośrodek czuciowy mowy (Wernickego) – tylna część górnego zakrętu skroniowego w lewej
półkuli. Jest to okolica pozwalająca identyfikować i różnicować fonemy ludzkiej mowy, czyli
po prostu pozwalająca rozumieć mowę.
Kora przedczołowa (prefrontal cortex) – kora płata czołowego leżąca najbardziej z przodu,
nie związana z funkcjami strikte ruchowymi płata czołowego i mową, obejmująca pola
Brodmanna 9, 46, 10, 47, 11, 12, 32. Jest to okolica związana z najbardziej złożonymi
procesami poznawczymi, takimi jak kontrola popędów, planowanie, monitorowanie przebiegu
zachowania i procesów poznawczych, ogólnie – jest związana z funkcjami wykonawczymi.
Kora oczodołowa (orbital cortex) – część kory przedczołowej leżącej na dolnej, brzusznej
części płata czołowego. Istnieje kilka powodów dla których można ją traktować oddzielnie,
jako składnik kory przedczołowej, ale też i kory mózgu w ogóle. Jest związana z kontrolą
emocjonalną, zdolnością do hamowania popędów i odraczania nagród. Jej temat będzie
jeszcze poruszony poniżej.

Hipokamp (hippocampus) – zawinięty do środka półkuli fragment kory płata skroniowego
na powierzchni przyśrodkowej, związany z procesami zapamiętywania nowych informacji w
zakresie pamięci deklaratywnej. Temat hipokampa będzie jeszcze poruszony poniżej.
Zakręt wrzecionowaty (fusiform gyrus) – znajduje się na powierzchni brzuszno-
przyśrodkowej płata skroniowego i będzie jeszcze o nim mowa poniżej. W obrębie tego
zakrętu lokalizuje się obszar związany z identyfikacją twarzy (fusiform face area, FFA).
Zakręt obręczy (cingulate gyrus) – przebiega na powierzchni przyśrodkowej łukowato nad
spoidłem wielkim, wchodząc w skład płata czołowego i ciemieniowego. Jest dzielony na
części przednią i tylną, z których każda ma nieco odmienne połączenia z innymi okolicami
mózgu. Przednia część zakrętu składa się z części grzbietowej – pole Brodmanna 32 i
brzusznej – pole Brodmanna 24. Część tylna składa się z części grzbietowych – pole
Brodmanna 31, oraz brzusznych – pole Brodmanna 23. W obrębie przednich części pól 23 i
31 niektórzy wydzielają jeszcze środkową część zakrętu obręczy. Każda z tych części
(przednia, środkowa i tylna) wykazuje nieco odmienne właściwości funkcjonalne.
Kilka dodatkowych słów opisu anatomicznego należy poświęcić na formację hipokampa.
Zaczniemy jednak od struktur jakie czytelnik miał już okazję poznać. Na bocznej powierzchni
płata skroniowego najniżej usytuowany jest dolny zakręt skroniowy. Jest on również
widoczny z perspektywy brzusznej, a także przyśrodkowej. Na powierzchni przyśrodkowej
nad nim znajduje się zakręt potyliczno-skroniowy boczny (lateral occipitotemporal gyrus),
nazywany również zakrętem wrzecionowatym (fusiform gyrus). Można przyjąć, że na mapie
Brodmanna pole 20 określaja dolny zakręt skroniowy, zaś na powierzchni przyśrodkowej pole
36 wskazuje zakręt wrzecionowaty. Kolejnym, ostatnim już zakrętem jest zakręt
przyhipokampowy (parahippocampal gyrus), mający w przednich częściach
charakterystyczną wypukłość – hak (uncuc). Tworzy on korę okołowęchową (perirhinal
cortex) – pole 35, oraz korę śródwęchową (entorhinal cortex) – pola 34 i 28. Kora
śródwęchowa przechodzi następnie w zawijającą się do wnętrza półkuli podkładkę,
utworzoną z trzech następujących po sobie części presubiculum (odpowiada jej pole 27),
subiculum i postsubiculum, po których następują już znacznie zawinięte do wnętrza półkuli
sektory Rogu Ammona (Ammon Horn; łac. Cornu Ammonis) CA1 – CA4. Ostatni z nich
kończy się przed łukowato biegnącym zakrętem zębatym (denate gyrus), który jakby zawija
się wokół zakończenia Rogu Ammona. Zakręt zębaty i cztery sektory Rogu Ammona tworzą
hipokamp (hippocampus; także hipokamp brzuszny, lub hipokamp właściwy). Struktura ta
cięgnie się od przednich części płata skroniowego do tylnych. Stamtąd wychodzi zawijając się
do góry nawleczka szara (indusium griseum; supracallosal gyrus), która przebiega następnie
do przodu nad spoidłem wielkim. Od tylnych części hipokampa odchodzi także pęk włókien
nerwowych, który również zawija się do góry, lecz następnie biegnie do przodu pod spoidłem
wielkim, zaś przed przegrodą przeźroczystą (septum pellucidum; błona rozdzielająca komory
boczne) zawija się w dół dochodząc do ciała suteczkowatego podwzgórza. Ta struktura
nazywana jest sklepieniem (fornix) i tworzy spoidło hipokampa, gdzie włókna z jednej
półkuli przechodzą do drugiej. W tym miejscu niezbędna wydaje się ilustracja: 1 | 2 | 3 | 4
| 5 .
Hipokamp jest utworzony z trzech nałożonych na sobie warstw korowych, określanych jako
kora stara archicortex. Wspomniana powyżej podkładka stanowi sferę przejściową między
korą starą a sześciowarstwową korą zakrętu przyhipokampowego. Trójwarstwowa kora
tworzy również pewne obszary ulokowane na korze oczodołowej, określane mianem kory
dawnej paleocortex. Kora stara i dawna są łącznie nazywane są korą niejednorodną

allocortex. Kora niejednorodna ma starszą historię ewolucyjną, aniżeli pozostała część kory
półkul mózgu, bywa też określana mianem kory węchowej. Na powierzchni kory
oczodołowej znajdują się struktury bezpośrednio związane ze zmysłem węchu: opuszki
węchowe (olfactory bulb), odbierające informacje zmysłu węchu i przekazujące je dalej do
kory za pomocą pasma węchowego (olfactory tract).
Należy tutaj podkreślić, że wytworzenie w toku ewolucji struktur przetwarzających
informacje węchowe nie miało na celu zapewnienia zwykłej percepcji różnych zapachów, ale
obwodów neuronalnych generujących prymitywne reprezentacje poznawcze otoczenia i
zapewniające bardziej adekwatne reakcje behawioralne. Na niższych szczeblach
ewolucyjnych zmysł węchu ma kluczowe znaczenie, pozwala różnicować obiekty,
rozpoznawać różne gatunki, różnych osobników w obrębie danego gatunku, jak i oceniać
potencjalne zagrożenie. Dla ludzi kluczowe znaczenie odgrywa zmysł wzroku, zaś zmysł
węchu został znacznie zredukowany. Jednak niektóre pierwotne aspekty częściowo zostały
zachowane i czytelnik dobrze je zna ze swojego subiektywnego doświadczenia. Kiedy tylko
poczujemy intensywną cuchnącą woń, ogarnia nas uczucie wstrętu i mamy ochotę czym
prędzej oddalić się od źródła nieprzyjemnego zapachu. Taki znacznie zautomatyzowany,
szybki i nie do końca świadomy sposób poznawania świata określamy mianem emocji. Zmysł
węchu miał na celu uruchamiać motywacje i określone sekwencje behawioralne, które
najprościej opisać w kategoriach zbliżania się do lub oddalania się od obiektu. Co więcej,
niższe organizmy mają dodatkowy układ węchowy przeznaczony na przetwarzanie informacji
dostarczanej przez feromony i związanej z aktywnością płciową. Tradycyjne koncepcje wiążą
rozwój węchomózgowia z tzw. środkową warstwą mózgowia (Yakovlev), mózgiem
paleossaków (MacLean), czy układem limbicznym (Papez). Kontynuując tę linię hipotez
można wyróżnić dwa obszary limbiczne, związane z obiema postaciami kory allocortex:
Oczodołowa strefa paralimbiczna: kora dawna paleocortex rozwija się do tyłu, na
powierzchni brzusznej i bocznej, do wyspy, bieguna płata skroniowego i przedniej części
kory przyhipokampowej, jak i w górę, przez przednie części zakrętu obręczy. Strefie tej
przypisuje się związki z interakcjami społecznymi, nastrojem, kontrolą emocjonalną oraz
ogólnie – z nieświadomym przetwarzaniem informacji.
Hipokampowa strefa paralimbiczna: kora stara archicortex rozwija się przez korę
śródwęchową do tylnych okolic zakrętu przyhipokampowego i dalej, na tylną część zakrętu
obręczy. Strefie tej przypisuje się związki z procesami pamięciowymi, motywacyjnymi,
uwagowymi, zaś ogólnie – ze świadomym przetwarzaniem informacji.
Tymczasem większość kory pokrywającej półkule mózgu po ich bocznych, grzbietowych,
brzusznych i przyśrodkowych częściach jest utworzona z powstałej w rozwoju ewolucyjnym
„niedawno” kory nowej neocortex, określanej też mianem kory jednorodnej isocortex,
utworzonej z sześciu warstw neuronalnych położonych jedna na drugiej. Kora nowa zajmuje
się „niższymi” funkcjami związanymi z odbiorem informacji zmysłowych i kontrolą
motoryczną, oraz funkcjami „wyższymi”, związanymi z językiem, mową, planowaniem,
organizowaniem, przewidywaniem konsekwencji i rozwiązywaniem problemów. Niektórzy są
skłonni stwierdzić, że to, co jest najbardziej ludzkie, znajduje się w korze nowej, która
najwyższe stadium ewolucji osiągnęła właśnie w naszych mózgach, zarówno w aspekcie
ilościowym, jak i jakościowym.
Jak już wspominano, kora nowa w mózgu człowieka utworzona jest przez sześć warstw.
Warstwa IV zajmuje się odbieraniem informacji spoza kory, zaś warstwa V zajmuje się

wysłaniem informacji poza korę. Komórki IV warstwy przekazują pobudzenia wyżej, do
warstw II i III, skąd informacja jest wysyłana do sąsiednich i innych okolic korowych, ale
również pobudza dendryty komórek warstwy V. Obszary sensoryczne odbierające informację
danej modalności zmysłowej ze wzgórza, mają znacznie rozbudowaną IV warstwę kory, która
można być dzielona na różne podwarstwy. Okolice ruchowe mają natomiast znacznie mniej
komórek w warstwie IV, ale za to dużą liczbę olbrzymich komórek piramidowych Betza w V
warstwie, których aksony tworzą następnie drogę korowo-rdzeniową. Oprócz kory
motorycznej M1, aksony do drogi korowo-rdzeniowej wysyłają także okolice przedruchowe
(leżące do przodu od M1) oraz kora somatosensoryczna. Tak więc zależnie od czynności,
jakie dana okolica pełni, mogą występować różnice w szczegółowej budowie warstw
korowych. Ze względu na subtelne różnice tej budowy tworzy się mapy okolic korowych, z
których najpopularniejszą jest mapa Brodmanna, o czym było już wcześniej wspomniane.
Głównym neuroprzekaźnikiem pobudzającym kory nowej jest kwas glutaminowy (Glu), zaś
hamującym – kwas gamma-aminomasłowy (GABA). Charakterystycznym neuronem jest
neuron piramidowy, mający charakter pobudzający. Znaczy to, że jego pobudzenie prowadzi
do wygenerowania potencjału czynnościowego, który uruchamia procesy wydzielania kwasu
glutaminowego na synapsach tworzonych przez zakończenia aksonalne. Aksony tych
komórek mogą być bardzo długie (jak w drodze korowo-rdzeniowej – od kory mózgu do
rdzenia kręgowego). Dochodzące do kory aksony ze wzgórza, oraz aksony pośredniczących
neuronów pobudzających tworzą swoje synapsy zazwyczaj z dendrytami komórek
piramidowych. Tymczasem komórki hamujące mają raczej charakter lokalny, tj.
pośredniczący, a za pomocą swych krótkich aksonów tworzą synapsy z ciałem komórkowym
neuronu piramidowego. Jest to ciekawe o tyle, że im bliżej ciała komórkowego znajdują się
synapsy, tym większy mają wpływ na generowanie pobudzenia. Jednostką funkcjonalną kory
nowej nie jest jednak pojedynczy neuron, lecz grupa neuronów wszystkich warstw tworząca
kolumnę korową (cortical column). W ten sposób kora mózgowa zapewnia skomplikowane
moduły przetwarzania informacji. Niezwykle trudno jest ustalić precyzyjne granice takich
kolumn. Jest to bowiem zależne od przyjętej metodologii. Ponieważ każda okolica kory
nowej jest silnie połączona ze wzgórzem, jako jednostkę podziału można przyjąć akson
wzgórza, który wnika do kory, a granicę kolumny wytyczyć po przez zakres tworzonych
przez niego synaps. Zarazem podobnie można uczynić z komórką piramidową V warstwy i
granicę kolumny określić na podstawie jej synaps. Można wreszcie określać odmienne
właściwości fizjologiczne, zwłaszcza różnice w odpowiedzi neuronów na bodźce i na tej
podstawie oznaczać kolumnę. Obraz kolumn wyznaczonych różnymi metodami jednakże nie
musi pokrywać się ze sobą. Dużo wiedzy na temat takich kolumn przyniosły badania
dotyczące pierwszorzędowej kory wzrokowej. Znajdują się tam kolumny reagujące na bodźce
o kształcie linii, gdzie każda kolumna przetwarza linie o różnym nachyleniu, zależnie też od
miejsca w polu wzrokowym, z jakiego pochodzi bodziec.
Jako struktury podkorowe tradycyjnie uznaje się jądra podstawy takie jak przedmurze, gałkę
bladą, skorupę i jądro ogoniaste, oraz ciało migdałowate. Ciało migdałowate (amygdala) jest
w istocie zespołem jąder migdałowatych, z których każde ma nieco odmienną charakterystykę
funkcjonalną i odmienne połączenia z innymi strukturami o.u.n. Jest związane z układem
autonomicznym emocjami, szczególnie zaś z lękiem. Znajduje się w przednich częściach
płata skroniowego, tuż przed hipokampem. Jest silnie powiązane z podstawnymi częściami
płata czołowego, z korą oczodołową i generalnie z oczodołową strefą paralimbiczną. Podczas
gdy ciało migdałowate tworzy stosunkowo „punktową” strukturę, przypominającą coś na
kształt kulki, tak pozostałe jądra podstawy znajdują się bardziej w centralnych rejonach
półkuli i są on bardziej „rozciągnięte”. Jądro ogoniaste ma chyba najbardziej

charakterystyczny kształt. Przednia część określana jest mianem głowy, od niej odchodzi do
tyłu ogon, mający łukowaty kształt zbliżony do górnej części komory bocznej i spoidła
wielkiego. Jądro ogoniaste wraz ze skorupą są określane łącznie mianem prążkowia
grzbietowego (lub po prostu – prążkowia).
Gałka blada i prążkowie grzbietowe (nazywane też prążkowiem nowym – neostriatum)
tworzą pętle neuronalne z korą motoryczną, przedruchową, a także z móżdżkiem, biorąc
udział w kontroli ruchów celowych i tych niezależnych od woli. Sieć neuronalna kontrolująca
ruchy niezależne od woli jest określana mianem układu pozapiramidowego (extrapyramidal).
Wielu autorów odchodzi obecnie od klasyfikacji czysto anatomicznej, i jako jądra podstawy
uznaje wszystkie podkorowe struktury mózgowia, które współtworzą układ pozapiramidowy.
Z tej klasyfikacji wyłącza się oczywiście ciało migdałowate, natomiast włącza się do niej
jądra niskowzgórza (międzymózgowie; nie były one wcześniej opisywane) oraz istotę czarną
(śródmózgowie). Informujemy o tym, aby czytelnik nie czuł się zaskoczony, jeżeli napotka
taką klasyfikację. Ostatnio pojawia się coraz więcej danych pokazujących, że gałka blada i
prążkowie są związane również z szeregiem innych procesów niż tylko z kontrolą
motoryczną. Zwraca się uwagę na udział prążkowia w automatyzowaniu czynności
dotychczas pozostających pod świadomą kontrolą korową, jak i przy nabywaniu nowych
procedur motorycznych i umiejętności (skill learning; pamięć proceduralna). Jądro ogoniaste,
skorupę i gałkę bladą można podzielić na różne części, które mają odmienne połączenia i
biorą względnie niezależny udział w różnych obwodach np. związanych z funkcjami
motywacyjnymi i przetwarzaniem informacji emocjonalnych w odpowiedź behawioralną
(brzuszne i przyśrodkowe części jądra ogoniastego i gałki bladej). Istnieją też prace
pokazujące na udział jąder podstawy w obwodach kontrolujących pamięć roboczą. Generalnie
w ostatnich latach znacznie więcej poświęca się uwagi udziałowi struktur podkorowych (w
tym i wzgórza) w funkcjach, jakie pierwotnie lokalizowano jedynie w korze.
W okolicy najbardziej przednich części głowy jądra ogoniastego i przegrody przeźroczystej
znajduje się jądro półleżące nucleus accumbens septi, leżące jakby na samym dnie półkul
mózgu. Określane jest jako prążkowie brzuszne, albo przynajmniej część prążkowia
brzusznego. Struktura ta jest związana z tzw. układem nagrody oraz procesami
motywacyjnymi, także tymi prowadzącymi do patologii związanej z motywacją (np.
uzależnienia). Wspólnie z ciałem suteczkowatym podwzgórza otrzymuje informacje z
hipokampa po przez sklepienie. Inną strukturą znajdującą się w przednich częściach
kresomózgowia jest jądro Meynerta. Stanowi ono ważny ośrodek rozsyłający po mózgowiu
aksony cholinergiczne, w tym i do hipokampa. Struktura ta ulega degeneracji w chorobie
Alzheimera, podobnie jak formacja hipokampa. Generalnie w bliskim sąsiedztwie przegrody
znajdują się tzw. jądra przegrody, z których część również wysyła do innych części mózgowia
aksony cholinergiczne podobnie jak jądro Meynerta.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}