Wpływ różnych czynników na szybkość reakcji enzymatycznych.
Wyjściowe stężenie substratu, enzymu
Czynniki zewnętrzne:
Temperatura
pH środowiska
Obecność aktywatorów i inhibitorów
Lepkość roztworu, skład roztworów, zanieczyszczenie itp.
Stężenie substratu, w jednakowych warunkach:
Przy małym stężeniu substratu szybkość reakcji chemicznej wzrasta wprost proporcjonalna do stężenia substratu. (Wzrost stężenia enzymu powoduje nieznacznie wzrost szybkości reakcji ale nie będzie to wzrost liniowy)
Przy średnim stężeniu substratu szybkość reakcji wzrasta, i jest równe KM, osiąga połowę Vmax.
Przy dużym stężeniu substratu, nie ma wpływu na szybkość reakcji, osiąga wartość maksymalną. Dochodzi do tego w momencie, gdy wszystkie cząsteczki enzymu są połączone z substratem, tworząc kompleksy E - S. (Wzrost stężenia enzymu powoduje wzrost wprost proporcjonalnie szybkości reakcji)
Temperatura.
Wzrost temperatury powoduje zwiększenie szybkości reakcji enzymatycznej, po osiągnięciu optimum szybkość ta zaczyna spadać ze względu na denaturację cieplną enzymu. Optymalna temperatura zależy od długości inkubacji (gdy enzym ma działać kilka sekund, może być wysoka, gdy kilka dni - musi być niska, by długo zachował aktywność), W temperaturze do ok. 37oC (w pH zbliżonym do punktu izoelektrycznego, temperatura powoduje szybszą denaturację białka, i utratę zdolności katalitycznej), podniesienie temp. o 10oC powoduje 2-krotne zwiększenie szybkości reakcji. Obniżanie temperatury zmniejsza szybkość reakcji biochemicznych, ale nawet zamrożenie enzymu nie powoduje trwałego utracenia jego aktywności; ponowne ogrzanie przywraca zdolność katalityczną enzymu.
Zmianę szybkości reakcji w zależności od temperatury według której wzrost temperatury o 10 0K powoduje 2-4-krotny wzrost szybkości reakcji. Z regułą van't Hoffa wiąże się temperaturowy współczynnik szybkości reakcji (TWSR), wyznaczający stosunek szybkości reakcji chemicznej po zmianie temperatury do szybkości reakcji przed zmianą temperatury (wzrostem lub spadkiem).
ΔT = T2 - T1 - zmiana temperatury.
γ - temperaturowy współczynnik szybkości reakcji (TWSR)
v1 - szybkość reakcji przed podwyższeniem temperatury
v2 - szybkość reakcji po podwyższeniem temperatury
pH. Szybkość reakcji jest optymalna w optymalnym pH, czyli większość w pH około 7, ale zależy to od rodzaju występowania enzymu, enzymy działające pozakomórkowo, w świetle przewodu pokarmowego, charakteryzują się znacznym zróżnicowaniem optymalnych warunków kwasowości środowiska. Wpływ pH na aktywność enzymów tłumaczy się tym, że są one białkami i są zależne od kwasowości środowiska i zjonizowanej formy, ogólne ramy określamy jako niskie i wysokie.
Inhibitory i aktywatory występujące jako jony metali ciężkich, które różnie działają w zależności od enzymów o które chodzi, mogą być dodane jak i ujemne, obecne jako zanieczyszczenia w odczynnikach, substancje niskocząsteczkowe, które przyłączając się do enzymu powodują zmianę struktury przestrzennej enzymu, uniemożliwiając tworzenie kompleksów E-S. Substancje te mogą również działać jako aktywatory, przyłączając się do centrum aktywnego i polepszenie wiązania substratu. Substancje te nie biorą udziału w reakcji, jednak aktywują lub zwiększają aktywność enzymów. Można je podzielić na trzy grupy: kofaktory, makrocząsteczki białkowe, drobnocząsteczkowe połączenia organiczne.
Energia aktywacji, reguła van't Hoffa, współczynnik temperaturowy.
Zmianę szybkości reakcji w zależności od temperatury według której wzrost temperatury o 10 0K powoduje 2-4-krotny wzrost szybkości reakcji. Z regułą van't Hoffa wiąże się temperaturowy współczynnik szybkości reakcji (TWSR), wyznaczający stosunek szybkości reakcji chemicznej po zmianie temperatury do szybkości reakcji przed zmianą temperatury (wzrostem lub spadkiem).
ΔT = T2 - T1 - zmiana temperatury.
γ - temperaturowy współczynnik szybkości reakcji (TWSR)
v1 - szybkość reakcji przed podwyższeniem temperatury
v2 - szybkość reakcji po podwyższeniem temperatury
Rodzaje inhibitorów, przykłady różnych typów, mechanizmy inhibicji enzymów.
Kationy aktywują niektóre enzymy (enzymy przenoszące elektrony - fosfatazy, dehydrogenazy, lipaza i inne) należą Mg2+, Zn2+, Mn2+, Ca2+, Cu2+, Fe2+, Fe3+, Co2+, Ni2+, Na+, K+.
Koenzymami są najczęściej witaminy i ich pochodne, odgrywają one rolę zwłaszcza w enzymach nieproteolitycznych. W przypadku enzymów proteolitycznych zasadnicze znaczenie mają jony Ca2+ (stabilizują aminopeptydazy, proteinazy serynowe i tiolowe), Co2+ (aktywują aminopeptydazy), Mn2+ (wpływają na aktywność prolidazy) i Zn2+ (warunkuje aktywność karboksypeptydazy A). Metale ciężkie hamują działanie enzymów.
Aniony wywierają niewielki wpływ na działanie enzymów, wyjątek stanowi aktywowanie amylazy oraz katepsyny C przez chlorki.
Niektóre enzymy wytwarzane są w postaci nieaktywnej, jako prekursory enzymów (proenzymy lub zymogen), a ich działanie pobudzane jest przez przekształcenie w wyniku aktywności innych enzymów, np. enterokinaza przekształca nieaktywny trypsynogen w aktywną trypsynę. Następuje to najczęściej przez przecięcie łańcucha peptydowego proenzymu w określonym miejscu, co powoduje zmianę konformacji peptydu i odsłonięcie miejsca aktywnego.
Aktywatorami nazywamy także czynniki, które usuwają wpływ inhibitorów, ograniczających czasowo działanie enzymu. Aktywność wielu enzymów jest hamowana w obecności łagodnych środków utleniających, np. tlenu atmosferycznego, co jest katalizowane przez metale ciężkie, znajdujące się w śladowych ilościach w odczynnikach lub materiale badawczy. Taka inhibicja jest często odwracalna, gdy np. zredukujemy grupę -S-S- za pomocą cysteiny, odtworzona zostanie aktywna grupa -SH, natomiast związki kompleksujące mogą związać metale ciężkie. Do enzymów, których aktywność zależy od obecności wolnych grup tiolowych należą katepsyny B, L i H.
Metody określenia typów inhibicji.
Inhibicja może być odwracalna i nieodwracalna, inhibicja odwracalna zaś może być kompetycyjna lub niekompetycyjna. Działanie inhibitora kompetycyjnego można rozpoznać po zastosowaniu równania Lineweavera-Burka - mierzy się szybkości początkowe v0 przy różnych stężeniach substratu i stałym stężeniu inhibitora.
Inhibicja kompetycyjna (współzawodniczący o miejsce aktywne) zmienia nachylenie prostej oraz punkt przecięcia z osią x (wzrasta KM), natomiast punkt przecięcia z osią y pozostaje bez zmian (stała wartość Vmax).
Inhibitora niekompetycyjnego (niewspółzawodniczący o miejsce aktywne) zwiększenie nachylenia prostej oraz zmianę punktu przecięcia z osią y (maleje Vmax), natomiast punkt przecięcia z osią x pozostaje bez zmian (KM pozostaje bez zmian).
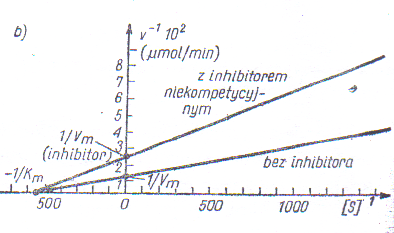
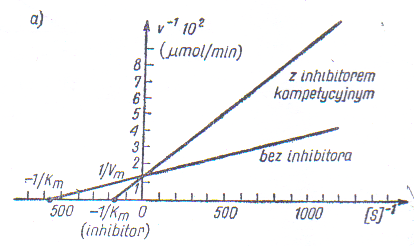
Przykłady inhibitorów i ich wpływu na reakcje enzymatyczne w technologii żywności.
Inwertaza jest enzymem katalizującym reakcję hydrolizy sacharozy do fruktozy i glukozy. W przemyśle spożywczym inwertaza jest wykorzystywana np. podczas produkcji czekoladek nadziewanych półpłynnym nadzieniem.
NaCl i mocznik reakcja enzymatyczna hydrolizy skrobi.
Chymotrypsyna pH 8,0 aktywowana przez trypsynę, powoduje powstawanie gorzkich peptydów.
Pepsyna pH 1-2 aktywowana przez jony H+ kwasu solnego W przemyśle spożywczym pepsyna wieprzowa i wołowa zastępują renninę przy produkcji serów.
Rennina (podpuszczka) pH 5,5-7,0 w obecności jonów Ca2+. Przekształca kazeinę w parakazeinian wapnia - ścina białka mleka, hydrolizuje także dalej frakcje kazeinowe w czasie dojrzewania sera,
Stałe kinetyczne, opisujące aktywność enzymów.
Stała Michaelisa - jest to taka stała która określa sprawność enzymu, jest to takie stężenie substratu, przy którym szybkość reakcji jest równa połowie szybkości maksymalnej.
Vmax - maksymalna szybkość
v - początkowa szybkość reakcji
[S] - molowe stężenie substratu
Szybkość maksymalna - Przy dużym stężeniu substratu, stężenie nie ma wpływu na szybkość reakcji i osiąga wartość maksymalną.
Właściwości i działanie enzymów amylolitycznych.
Amylaza to ogólna nazwa enzymów hydrolizujących skrobię, Optymalne warunki działania amylaz to, w zależności od pochodzenia enzymu, pH lekko kwaśne (zbożowe 50-65oC) do obojętnego (zwierzęce 37oC), w obecność jonów Cl-. do których zaliczamy;
α-amylazę, endoamylazę rozkładającą skrobię od środka wiązania α-1,4-glikozydowe, nie działa na rozgałęzienia; powstają α-dekstryny o małej masie cząsteczkowej i niewielka ilość maltozy.
β-amylazę, rozkładającą skrobię od końca łańcucha, rozcinając wiązania α-1,4-glikozydowe oraz amyloglukozydazę, powstają duże amylodekstryny i znaczna ilość maltozy i dekstryna graniczna
glukoamylazę, odcinające glukozę na końcu łańcucha skrobiowego po jednej cząsteczce od zewnątrz i powstaje glukoza, rozkładając zarówno wiązania α-1,4-glikozydowe, jak i α-1,6-glikozydowe.
Działanie wszystkich enzymów może zhydrolizować skrobię do glukozy.
Mechanizmy aktywacji i inhibicji enzymów.
Pod wpływem mikroelementów, np. jonów metali, kofaktorów, makrocząsteczek białkowych, drobnocząsteczkowych połączeń organicznych, może nastąpić taka zmiana kształtu cząsteczki enzymu, która jest korzystna dla przebiegu reakcji enzymatycznej. Odbywa się to na skutek przyłączenia aktywatora do centrum aktywnego i polepszenia w ten sposób wiązania substratu. Enzym i substrat ma komplementarne miejsce do połączenia się i powstaje kompleks ES, aby przyśpieszyć reakcje dodaje się różnego typu aktywatory które aktywują lub zwiększają aktywność enzymów.
Enzym może wiązać albo cząsteczkę substratu, albo cząsteczkę inhibitora ale nie obie równocześnie. W zależności od tego czego będzie więcej reakcja będzie przebiegała wolniej lub szybciej, jeśli będzie więcej inhibitora reakcja będzie przebiegała wolniej i odwrotnie. Po reakcji enzym jest uwalniany w tej samej postaci i może wziąć udział w innej reakcji. Aktywatorami nazywamy także czynniki, które usuwają wpływ inhibitorów, ograniczających czasowo działanie enzymu.
Aktywność enzymów może być zakłócana przez działanie wielu substancji. Katalizatory ujemne, zmniejszają szybkość reakcji nazywamy inhibitorami. Inhibicja może być odwracalna i nieodwracalna, inhibicja odwracalna zaś może być kompetycyjna lub niekompetycyjna.
Rodzaje substancji aktywujących, ich wpływ na działanie odpowiednich enzymów.
Aktywatory - substancje te nie biorą udziału w reakcji, jednak aktywują lub zwiększają aktywność enzymów. Można je podzielić na trzy grupy:
1) kofaktory - koenzymy, grupy prostetyczne oraz jony niektórych metali, wbudowanych w cząsteczkę apoenzymu, a także niektóre aniony nieorganiczne
2) makrocząsteczki białkowe, odkrywające grupy czynne enzymu, przekształcające nieaktywną formę enzymu w aktywną
3) drobnocząsteczkowe połączenia organiczne, usuwające wpływ związków hamujących, redukujące potencjał oksydoredukcyjny środowiska
Inwertaza i oksydaza glukozowa. trypsyna w pH 1 i 90 0 C
Inwertaza (E.C. 3.2.1.26) jest wytwarzana przez komórki drożdży, jest enzymem katalizującym reakcję hydrolizy sacharozy, do glukozy i fruktozy. W przemyśle spożywczym inwertaza jest wykorzystywana np. podczas produkcji czekoladek nadziewanych półpłynnym nadzieniem. Podczas produkcji nadzienie w postaci stałej (głównym składnikiem jest sacharoza) pokrywane jest czekoladą, a następnie w ciągu kilkutygodniowego „dojrzewania“ czekoladek w wyniku działania inwertazy dodanej w procesie produkcji, sacharoza rozkładana jest do fruktozy i glukozy, które chłonąc wilgoć ze środowiska powodują w efekcie upłynnienie nadzienia.
Oksydaza glukozowa utlenia glukozę do kwasu glukozowego. Enzym ten wykazuje powinowactwo do β-D-glukopiranozy. Oksydazę glukozową stosuje się do ilościowego oznaczania glukozy w żywności, we krwi i w moczu, a także do usuwania glukozy z produktów spożywczych oraz do usuwania tlenu z puszek z konserwami, piwem czy też z opakowań foliowych.
Izolacja i oczyszczanie enzymów.
Enzymy są przeważnie bardzo nietrwałe w środowisku o pH mniejszym niż 5 lub większym niż 9, łatwo ulegają też denaturacji cieplnej (powierzchniowej - podczas pienienia się roztworów) oraz są inaktywowane przez metale ciężkie.
Izolowanie należy przeprowadzać w taki sposób, aby zapobiegać utracie zdolności katalitycznych, tzn.
w buforach o pH około 7,
w niskiej temperaturze (najczęściej około 0oC),
stosując wodę podwójnie destylowaną oraz
środki kompleksujące metale ciężkie, a w razie konieczności również związki tiulowe,
zachowując wyjątkową czystość na stanowisku pracy.
Oczyszczanie enzymów
Dla każdego enzymu powinno się zastosować indywidualny schemat oczyszczania wykorzystujący jego własności fizykochemiczne.
1 etap oczyszczania białek jest przeprowadzenie ich do roztworu, chyba że izolujemy je z płynnego źródła (np. z krwi, śliny itp). Stosuje się w tym celu techniki doprowadzające do zniszczenia ścian, błon komórkowych - czynniki biologiczne - lizozym, ciśnienie enzymatyczne (sól, cukier), ultradźwięki, homogenizacja, rozrywanie komórek piaskiem lub perełkami szklanymi, inne metody mechaniczne,
2 etap to wstępne procesy oczyszczania: Dalsze metody stosowane w celu otrzymania preparatu enzymatycznego są bardzo różnorodne i ich stosowanie zależy od aktualnego i pożądanego stopnia oczyszczenia enzymu. Najczęściej stosowanymi wstępnymi procesami oczyszczania są:
- frakcjonowanie roztworami soli (np. siarczanem amonu)
- ekstrakcja rozpuszczalnikami organicznymi (np. etanolem, acetonem lub eterem).
3 etap - oczyszczanie Jako kolejny etap otrzymywania enzymów, szerokie zastosowanie ma obecnie chromatografia, elektroforeza, ultrawirowanie, stosowana jest także krystalizacja.
4 etap kontrola; Kontrola jakościowa (wzrastająca czystość białka), techniki elektroforetyczne (zwłaszcza SDS-PAGE) Kontrola ilościowa (stężenie interesującego enzymu i/lub jego aktywności), techniki spektrofotometryczne, np. metoda Lowry'ego, metoda biuretowa.
Enzymy proteolityczne, amylolityczne, lipolityczne.
Proteolityczne - rozkładają białka, Należą do nich: trypsyna, chymotrypsyna, karboksypeptydazy A i B, elastaza, nukleaza.
Amylolityczne - rozkładają złożone cukry. czyli trawiące skrobię. Jest to alfa-amylaza trzustkowa.
Lipolityczne - rozkładają tłuszcze. Należą do nich: lipaza trzustkowa, fosfolipazy A i B, esterazy.
Nukleityczne - rozkładają kwasy nukleinowe.
16. Metody badań aktywności enzymów.
Oznaczenie aktywności enzymów wykonuje się w oparciu o ich zdolności katalityczne. Stosuje się metodę Ansona, która opiera się na procesie hydrolizy zdenaturowanej hemoglobiny jako substratu. Na podstawie różnicy ilości produktów hydrolizy białka (PBH) w przesączach, otrzymanych po strąceniu roztworów 10% TCA, określa się aktywność proteolityczną badanego enzymu.
17. Jednostki aktywności enzymów.
Jednostka U każdego enzymu jest taka jego ilość która katalizuje przemianę 1µmola substratu w ciągu 1 min. W temp.30°C i w optymalnych warunkach. Ze względu na rząd różnic między aktywnością enzymów stosuje się przedrostki metryczne. Obecnie Międzynarodowa Unia Biochemiczna zaleca stosowanie nowej jednostki 1 katal. Odpowiada ona aktyw enzymu, przy której jest on zdolny przekształcić 1 mol substratu w produkt w czasie 1s w temp 30°C i pozostałych warunkach optymalnych. 1 katal = 6*107 U. 1U = 16,67nKat
Metody oznaczania stężenia białka i produktów hydrolizy białka.
Badanie ilości produktów hydrolizy białka prowadzi się najczęściej metodą Lowry'ego. Jest to metoda kolorymetryczna, w której końcowa barwa jest wynikiem po pierwsze, reakcji biuretowej białka z jonami miedzi w środowisku zasadowym, czyli przyłączenia jonów miedzi do wiązań peptydowych, po drugie, redukcji odczynnika fosfomolibdeno-fosfowolframowego (odczynnik Folina-Ciocalteu) przez obecne w białku tyrozynę i tryptofan (redukcję barwy powodują także związane już jony miedzi, natomiast barwa powstająca przy nieobecności jonów miedzi pochodzi tylko od aminokwasów).
Oznaczanie stężenia białka metodą biuretową: Do kolbek odmierzyć po 0,5 cm3 kolejnych prób (do próby ślepej użyć wody destylowanej), dodać po 2 cm3 odczynnika miedziowego (wszystkie próby w 3 powtórzeniach). Po 30 min odczytać absorbancję przy długości fali 540 nm. Zawartość białka odczytać z krzywej wzorcowej (po uwzględnieniu zmętnienia), następnie zestawić wyniki, biorąc pod uwagę rozcieńczenie.
Pytania które były:
Wymienić czynniki które wpływają na szybkość reakcji enzymatycznych.
Opisać wpływ temperatury.
Właściwości α-amylazy.
Metody określenia typów inhibicji i jak możemy cofnąć działanie inhibitora opisać każdą z osobna.
5
5
Wyszukiwarka
Podobne podstrony:
Świnie ćw kolo nr 3
Świnie ćw kolo nr 3
PCR RAPD Genetyka molekularna ćw koło 3
GN cw kolo 1 zagadnienia, Geodezja, Gospodarka nieruchomosciami, Materialy
A cw 2 koło 1 kinetyka reakcji zadania dodatkowe 2
Ćw 5 koło 2
analityczna-cw-kolo
mnja cw kolo sciaga do druku
A cw 6 koło 2
SCHORZENIA TRAWIEŃCA - ćw, I koło
Ćw 8 koło 2
A cw koło 2 mała
Ćw 7 koło 2
Ekonomia ćw kolo
Ćw 6 koło 2
Świnie ćw kolo nr 3
cw 5 - trucizny poch.roslinnego, Toksykologia, toksyki na 2 kolo
ćw żelbet kolo zaliczeniowe
cw 8 - preparaty stos. w gosp.domowym, Toksykologia, toksyki na 2 kolo
więcej podobnych podstron