
32
ÂWIAT NAUKI WRZESIE¡ 2004
Skarb
ukryty
w genomach
zbó˝
Krzy˝owanie roÊlin oparte
na znajomoÊci ich genomów
zapoczàtkuje
nowà zielonà rewolucj´
STEPEN A. GOFF
i JOHN M. SALMERON

WRZESIE¡ 2004 ÂWIAT NAUKI
33
SIEWKI RY˚U mo˝na testowaç genetycznie
na obecnoÊç po˝àdanych cech.

Przez tysiàce lat
rolnicy uprawiali pola, obserwujàc
niebo z nadziejà na dobrà pogod´ i ob-
fite zbiory. Kiedy znajdowali roÊliny,
które pomimo kiepskiej aury dawa∏y do-
bry plon i na dodatek przetrwa∏y zara-
zy, próbowali utrwaliç ich przydatne ce-
chy przez krzy˝owanie. Zawsze jednak
by∏o to dzia∏anie na chybi∏ trafi∏, gdy˝
nie mieli mo˝liwoÊci wglàdu do wn´-
trza roÊlin i nie wiedzieli, co decyduje
o ich w∏aÊciwoÊciach.
Pomimo elementu przypadkowoÊci
metoda ta dawa∏a znakomite rezultaty.
Kiedy oko∏o 10 tys. lat temu nasi przod-
kowie porzucili ∏owiectwo i zbieractwo
na rzecz rolnictwa, nastàpi∏a eksplozja
demograficzna, która trwa do dziÊ, wy-
muszajàc post´p w produkcji rolnej.
Obecnie 99% tej produkcji opiera
si´ na zaledwie 24 gatunkach roÊlin
uprawnych. Trzy z nich – ry˝, pszenica
i kukurydza – dostarczajà wi´kszoÊci
spo˝ywanych na Êwiecie kalorii, sà pro-
dukowane w iloÊci ponad pó∏ miliarda
ton rocznie ka˝dy, a ich Êrednie dzienne
spo˝ycie waha si´ od 0.4 do 1.5 kg na
osob´. Aby zaspokoiç zapotrzebowanie
ludzkoÊci, która przed 2050 rokiem
ma liczyç 9 mld osób, uprawy zbó˝ po-
winny dostarczaç co roku o 1.5% wi´-
cej pokarmu mimo systematycznego
zmniejszania si´ area∏u upraw.
Mistrzowska selekcja
NAUKOWCY
zajmujàcy si´ hodowlà i do-
borem odmian roÊlin uprawnych sà
przekonani, ˝e plony zbó˝ nie osiàgn´-
∏y jeszcze teoretycznie mo˝liwego pu-
∏apu i pracujà nad ich zwi´kszeniem.
Pojawienie si´ nowych narz´dzi badaw-
czych sprawi∏o, ˝e kluczem do sukcesu
mo˝e si´ okazaç analiza pochodzenia
trzech najwa˝niejszych zbó˝.
Badania molekularne i genetyczne
wskazujà, ˝e pszenica, ry˝ i kukurydza,
a tak˝e j´czmieƒ, proso, sorgo oraz in-
ne trawy sà znacznie bli˝ej spokrewnio-
ne, ni˝ wczeÊniej uwa˝ano. Poznanie
kodu genetycznego ka˝dego z tych ga-
tunków w du˝ym stopniu u∏atwia udo-
skonalenie pozosta∏ych. Wiele takich
ulepszeƒ mo˝e wynikaç z wykorzysta-
nia genetycznego bogactwa bezpo-
Êrednich przodków naszych zbó˝. Od-
powiednie krzy˝owanie pozwala na
odzyskanie wielu cennych cech utraco-
nych przez roÊliny uprawne podczas ty-
si´cy lat selekcji.
Wszystkie nasze zbo˝a sà potomkami
pospolitych traw, jednak ich poszcze-
gólne linie oddzieli∏y si´ od siebie oko-
∏o 50–70 mln lat temu, zasiedlajàc od-
r´bne regiony Êwiata. Uwa˝a si´, ˝e
blisko 10 tys. lat temu na obszarze
˚yznego Pó∏ksi´˝yca, rozciàgajàcym si´
od wybrze˝y Morza Âródziemnego po
Mezopotami´ i pó∏nocne kraƒce Syrii,
udomowiono pszenic´. JakieÊ 1000 lat
póêniej na obszarze obecnego Meksyku
tamtejsi rolnicy zacz´li uprawiaç przod-
ka wspó∏czesnej kukurydzy, natomiast
ponad 8 tys. lat temu Chiƒczycy opano-
wali upraw´ ry˝u.
Od momentu udomowienia tych ro-
Êlin nasi przodkowie systematycznie
zwi´kszali ich plonowanie metodà bar-
dzo zbli˝onà do nowoczesnej selekcji
odmian. Wychodzàc od dzikich gatun-
ków, wybierali, krzy˝owali i uprawiali
roÊliny majàce takie przydatne cechy,
jak wi´ksza liczba lub rozmiary ziaren
1
.
Atrakcyjne by∏y tak˝e roÊliny, które nie
wysiewa∏y ziaren, poniewa˝ ich zbiór
by∏ prostszy (choç roÊlina taka mia∏a
gorszà zdolnoÊç do samodzielnego roz-
przestrzeniania si´). Selekcjà kierowa∏a
równie˝ przydatnoÊç kulinarna. Po˝à-
dane by∏y na przyk∏ad ziarna z cienki-
mi delikatnymi ∏upinami, ∏atwiejsze do
zjedzenia, albo kukurydza o zawartoÊci
skrobi pozwalajàcej na przygotowanie
tortilli. Tym sposobem roÊliny uprawne
coraz bardziej ró˝ni∏y si´ od dziko ro-
snàcych i coraz rzadziej by∏y z nimi krzy-
˝owane. Wspó∏czesna kukurydza tak
bardzo odbiega wyglàdem od swego
przodka teosinte, ˝e do niedawna jej
prawdziwe pochodzenie powszechnie
kwestionowano [ilustracja powy˝ej].
Przekszta∏canie roÊlin uprawnych w
procesie selektywnej hodowli i krzy˝o-
wania trwa do dziÊ. W ubieg∏ym wieku,
aby uzyskaç wi´ksze plony, roÊliny zbo˝o-
we selekcjonowano pod kàtem wi´kszych
k∏osów. Takie pe∏niejsze k∏osy by∏y ci´˝-
sze, wi´c w przypadku ry˝u i pszenicy
wybierano ni˝sze roÊliny, aby nie pok∏a-
da∏y si´ przy silniejszym wietrze. Hodow-
la w kierunku odpornoÊci na choroby, to-
lerancji na stresy Êrodowiskowe, lepszego
przyswajania i wykorzystania nawozów
azotowych radykalnie podnios∏a plono-
wanie i jego regularnoÊç, dajàc poczàtek
zielonej rewolucji lat szeÊçdziesiàtych.
34
ÂWIAT NAUKI WRZESIE¡ 2004
KA
Y CHERNUSH (
popr
zednie str
ony
); NINA FINKEL (
rysunki na tej str
onie
); JOHN DOEBLEY (z
dj´cia na tej str
onie
)
n
Porównanie genomów g∏ównych roÊlin uprawnych wykaza∏o ich bliskie pokrewieƒstwo
oraz wk∏ad cz∏owieka w ukierunkowanie ich ewolucji.
n
Dzi´ki ustaleniu funkcji poszczególnych genów naukowcy mogà poszukiwaç wersji genów
warunkujàcych po˝àdane cechy poÊród roÊlin uprawnych i ich dzikich krewnych.
n
Wykorzystanie korzystnego genu jako ∏atwo wykrywalnego znacznika zwi´ksza szybkoÊç
i precyzj´ tradycyjnego krzy˝owania roÊlin.
Przeglàd /
Bezcenna ró˝norodnoÊç genetyczna
WSPÓ¸CZESNA KUKURYDZA I JEJ PRZODEK TEOSINTE wyglàdajà tak ró˝nie (rysunki powy˝ej),
˝e ich pokrewieƒstwo podawano w wàtpliwoÊç, dopóki nie potwierdzi∏y go badania genetyczne.
Dzi´ki selektywnej hodowli roÊlin charakteryzujàcych si´ po˝àdanymi cechami pradawni rolnicy
z obszaru obecnego Meksyku bezwiednie wybierali pewne wersje genów kontrolujàcych wzorzec
rozga∏´ziania p´du, struktur´ ziarna i inne cechy. Przed oko∏o 4400 laty wysmuk∏à, twardà kol-
b´ teosinte (zdj´cie z lewej) zastàpi∏a wspó∏czesna kolba kukurydzy (zdj´cie z prawej) i ju˝ wów-
czas warianty genów kontrolujàcych magazynowanie bia∏ek i jakoÊç skrobi by∏y takie same jak we
wszystkich wspó∏czesnych odmianach.
Teosinte Udomowiona
Teosinte
Wczesna
udomowiona
kukurydza kukurydza

Przyk∏adowo od 1950 roku w Stanach
Zjednoczonych Êredni plon kukurydzy z
hektara wzrós∏ o ponad 400%.
Jednak metody, jakimi pos∏ugiwali si´
hodowcy w tych latach, niewiele ró˝ni∏y
si´ od stosowanych w czasach pre-
historycznych. Wi´kszoÊç z nich, wy-
bierajàc roÊliny do dalszego rozmna˝a-
nia, wykorzystywa∏a dobrze widoczne
zewn´trzne cechy, takie jak wielkoÊç zia-
ren lub budowa p´du.
Badania genomów zbó˝ pokazujà jed-
nak, ˝e dawni rolnicy, selekcjonujàc ce-
chy zewn´trzne, w istocie nieÊwiado-
mie selekcjonowali poszczególne geny.
Zespó∏ Svante Pääbo z Max-Planck-In-
stitut für evolutionäre Anthropologie w
Lipsku w Niemczech analizowa∏ allele
(warianty) wybranych genów w kolbach
kukurydzy z wykopalisk z rejonów udo-
mowienia kukurydzy w Meksyku. Pääbo
i jego wspó∏pracownicy ustalili, ˝e ju˝
4400 lat temu uprawna kukurydza za-
wiera∏a takie same allele genów odpo-
wiedzialnych za wzorzec rozga∏´ziania
p´du oraz jakoÊç bia∏ka i skrobi zawar-
tej w roÊlinie, jakie odnajdujemy we
wszystkich wspó∏czesnych odmianach.
Natomiast u teosinte allele te wyst´pu-
jà zaledwie u 7–36% osobników, co ilu-
struje szybkie efekty silnej presji selek-
cyjnej, jakà wywarli pierwotni rolnicy.
Hodowcy, pracujàc niezale˝nie nad
ró˝nymi gatunkami zbó˝, zupe∏nie nie-
Êwiadomie zmieniali je przez kierunko-
wà selekcj´ mutacji zachodzàcych w po-
dobnych grupach genów. Mapowanie
cech – czyli przypisywanie przypusz-
czalnej lokalizacji genu warunkujàcego
okreÊlonà cech´ do konkretnego obsza-
ru chromosomu, tzw. locus – pokaza∏o,
˝e wiele cech wprowadzonych do wspó∏-
czesnych roÊlin uprawnych uda∏o si´ w
poszczególnych odmianach i gatunkach
zmapowaç w tych samych loci. A wi´c
mimo milionów lat niezale˝nej ewolu-
cji, dzielàcych poszczególne gatunki ro-
Êlin zbo˝owych, ich struktura genetycz-
na pozosta∏a nadal bardzo zbli˝ona.
Kilka tysi´cy loci warunkujàcych po-
szczególne cechy zosta∏o ju˝ zma-
powanych w ró˝nych udomowionych
roÊlinach. Stwierdzono przy tym zaska-
kujàco wysoki poziom zachowania mi´-
dzygatunkowego podobieƒstwa map ge-
netycznych tych roÊlin. Ten w∏aÊnie
wysoki poziom podobieƒstwa (nazywa-
ny te˝ syntenià), mi´dzy genomami traw
pozwoli∏ naukowcom uznaç te roÊliny
za jeden system genetyczny. Oznacza to,
˝e ka˝de odkrycie genu lub jego funkcji
w jednym z gatunków zbó˝ mo˝e po-
móc w identyfikacji genów lub udosko-
naleniu innego gatunku.
Wszystko wskazuje na to, ˝e pierw-
szym dobrze poznanym zbo˝em b´dzie
ry˝ (Oryza sativa). Ju˝ niebawem pozna-
my ca∏y jego genom. Jeden z nas (Goff)
opublikowa∏ ostatnio wst´pnà sekwen-
cj´ genomu podgatunku japonica, ry˝u
powszechnie uprawianego w Japonii i
Stanach Zjednoczonych. Naukowcy
chiƒscy ukoƒczyli sekwencjonowanie ge-
nomu podgatunku indica, uprawianego
w Azji kontynentalnej. Oczekuje si´, ˝e w
ramach Mi´dzynarodowego Projektu Po-
znania Genomu Ry˝u do koƒca tego ro-
ku uda si´ opracowaç szczegó∏owà se-
kwencj´ 12 chromosomów.
Uprawa genomów
GENOM RY
˚U
jest spoÊród genomów
wszystkich zbó˝ naj∏atwiejszy do zba-
dania. Zawiera zaledwie 430 mln par
nukleotydów DNA. Dla porównania
ludzki genom ma ich 3 mld. Genom
j´czmienia zawiera 5 mld par, natomiast
pszenica a˝ 16 mld. Projekt sekwen-
cjonowania genomu kukurydzy jest
obecnie realizowany, a sekwencjonowa-
nie pszenicy jest rozwa˝ane. Na pod-
stawie danych dotyczàcych sekwencji
genomu ry˝u zidentyfikowano ju˝ dzie-
siàtki tysi´cy konkretnych genów. Jed-
nak sama informacja, ˝e dany obszar
w genomie jest genem, nie mówi nieste-
ty nic o jego funkcji.
Istnieje wiele strategii pozwalajàcych
ustaliç funkcj´ nowego genu. Najprost-
sza polega na przeszukaniu bazy ziden-
tyfikowanych ju˝ genów w celu odnale-
zienia najbardziej podobnych. Wiele
genów odpowiada za tak podstawowe
procesy komórkowe, ˝e niemal identycz-
ne geny mo˝na znaleêç w poznanych
ju˝ genomach drobnoustrojów lub in-
nych wczeÊniej zbadanych organizmów.
SpoÊród 30–50 tys. genów (na tyle sza-
cowany jest genom ry˝u) oko∏o 20 tys.
ma prawdopodobnie sekwencj´ analo-
gicznà do genów ju˝ odkrytych, których
funkcje zosta∏y poznane. Pozwoli to na-
ukowcom z góry przewidzieç ich rol´.
Przypuszcza si´ na przyk∏ad, ˝e po-
nad tysiàc genów ry˝u jest zaanga˝o-
wanych w obron´ tej roÊliny przed pa-
togenami i szkodnikami. Podobnie setki
genów przyporzàdkowano do specyficz-
nych szlaków syntezy witamin, w´glo-
wodanów, lipidów, bia∏ek i innych po-
˝àdanych substancji. Na podstawie
danych z badaƒ nad dobrze ju˝ pozna-
nymi roÊlinami, takimi jak Arabidopsis
(rzodkiewnik pospolity), uda∏o si´ ziden-
tyfikowaç wiele genów regulujàcych
szlaki biosyntezy lub wp∏ywajàcych na
wa˝ne etapy rozwoju zbó˝, takie jak two-
rzenie kwiatów i rozwój nasion.
Wiele zespo∏ów naukowych posz∏o da-
lej i za pomocà mikromacierzy (chipów)
WRZESIE¡ 2004 ÂWIAT NAUKI
35
DOUG WILSON
Corbis
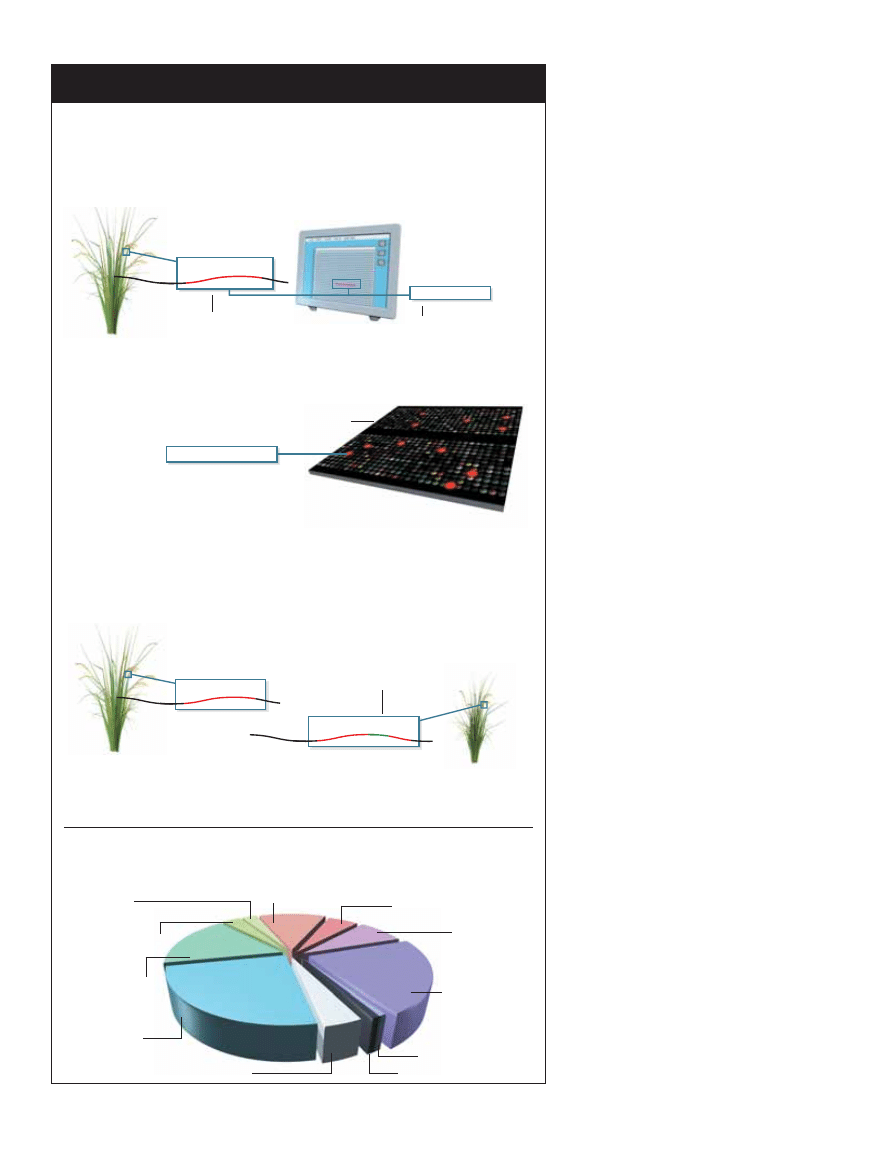
PO˚ÑDANE PRZEZ HODOWCÓW CECHY mo˝na ogólnie podzieliç na dotyczàce wzrostu,
budowy roÊliny, odpornoÊci na niekorzystne dzia∏anie Êrodowiska i zawartoÊci substan-
cji od˝ywczych. Zwi´kszenie plonów – marzenie rolników – mo˝na osiàgnàç, zwi´ksza-
jàc liczb´ lub mas´ ziaren pojedynczej roÊliny, zag´szczenie roÊlin lub ich tolerancj´ na
warunki, które wczeÊniej by∏y wysoce szkodliwe.
KRYTERIA DOBORU
Wzrost
Masa i liczba ziaren
Rozmiar k∏osa
SzybkoÊç
dojrzewania
Budowa
WysokoÊç
Rozga∏´zienie
Budowa kwiatu
OdpornoÊç na stres
Susza
Szkodniki
Choroby
Herbicydy
Intensywne nawo˝enie
ZawartoÊç/jakoÊç
sk∏adników od˝ywczych
Skrobia
Bia∏ka
T∏uszcze
Witaminy
DNA zacz´∏o katalogowaç geny, które sà
wyra˝ane (aktywowane) w poszczegól-
nych tkankach. Na przyk∏ad naukowcy
z naszej firmy Syngenta zbadali 21 tys.
genów ry˝u i zidentyfikowali 269 takich,
które sà szczególnie aktywne w fazie
powstawania ziaren, co sugeruje, ˝e od-
grywajà kluczowà rol´ w kszta∏towaniu
ich sk∏adu.
Nieco innym podejÊciem do identyfika-
cji funkcji genu jest jego unieczynnienie
(nokaut) poprzez wywo∏anie takiej muta-
cji w genie, która blokuje jego aktywnoÊç.
Nast´pnie obserwuje si´ skutki – czasem
efekt jest oczywisty, ale roÊlina mo˝e byç
tak˝e testowana pod kàtem mniej jedno-
znacznych zmian w przebiegu typowych
funkcji fizjologicznych, rozwojowych, bio-
chemicznych albo te˝ regulacji wewn´trz-
nej. Dzi´ki prywatnym i publicznym fun-
duszom zgromadzono ca∏e kolekcje
zmutowanych roÊlin ry˝u i kukurydzy, w
których zosta∏y w ten sposób zmienione
tysiàce specyficznych genów. Takie czyn-
noÊciowe badania genomu, ∏àcznie z po-
równywaniem sekwencji genów mi´dzy
gatunkami, pozwalajà naukowcom uzys-
kaç odpowiedzi na pytanie: ile i które z
genów ry˝u i – rozszerzajàc pytanie na in-
ne gatunki – które z genów kukurydzy,
pszenicy, sorgo i innych zbó˝ biorà udzia∏
w rozwoju roÊlin, sà odpowiedzialne za
fizjologi´, metabolizm i plonowanie.
Kiedy funkcja genu jest ustalona, wy-
korzystanie tej wiedzy w celu udoskonale-
nia zbo˝a wymaga identyfikacji poszcze-
gólnych alleli genu, które odpowiadajà za
po˝àdanà cech´. Na przyk∏ad jeÊli wia-
domo, ˝e dany gen kontroluje proces
gromadzenia skrobi w ziarnach kukury-
dzy, mo˝na poszukaç takiej wersji genu,
która funkcjonuje prawid∏owo mimo do-
tkliwej suszy. Takie po˝àdane allele genu
mogà zostaç odnalezione w innych wspó∏-
czesnych odmianach kukurydzy albo – co
bardziej prawdopodobne – wÊród dzikich
krewnych roÊlin zbo˝owych. Genetyczna
jednorodnoÊç wspó∏czesnych zbó˝ jest nie-
korzystnà konsekwencjà sposobu, w jaki
nasi przodkowie kiedyÊ je udomowili. We-
d∏ug jednego z szacunków wspó∏czesna
kukurydza powsta∏a z populacji roÊlin sk∏a-
dajàcej si´ z 20 osobników. Wybierajàc za-
ledwie kilka osobników majàcych cenne
cechy, a nast´pnie wzajemnie krzy˝ujàc
ich potomstwo przez tysiàce lat, ludzie
znacznie ograniczyli genetycznà ró˝no-
rodnoÊç roÊlin uprawnych.
Steven Tanksley i Susan R. McCouch,
naukowcy z Cornell University, badajàc
36
ÂWIAT NAUKI WRZESIE¡ 2004
SLIM FILMS
PRZYPISANIE GENÓW DO CECH
I I I I I I I I I
I I I I
I I I I I I
I I I
T C A G G ACCC
T G G A C
C T T G C
T C A
T G G A C C T T G C
A A T G C T A A C C
G G A C C G T T A
T G G A C C T A G C
T G G A C C T T G C
I I I I I I I I I
I I I I
I I I I I I
I I I
T C A G G ACCC
T G G A C
C T T G C
T C A
I I I I I I I I I
I I I I
I I I I
I I I
I I
I I I
T C A G G A C CC
T G G A C
C T T
T G A
G C
T C A
NARZ¢DZIA DO IDENTYFIKACJI genów ludzkich chorób nadajà si´ tak˝e do poszuki-
wania genów warunkujàcych poszczególne cechy roÊlin. Przypuszczalnà lokalizacj´ ge-
nu danej cechy za pomocà tzw. mapowania zaw´˝a si´ do konkretnego regionu w chro-
mosomie, a nast´pnie sekwencjonuje DNA tego regionu, by dok∏adnie wskazaç
prawdopodobny gen. Do ustalenia funkcji genu mo˝na zastosowaç nast´pujàce metody:
ANALIZY BAZ DANYCH
PROFILE EKSPRESJI GENÓW
KOLEKCJE MUTANTÓW
Odnaleziony gen porównuje si´ ze znanymi ju˝ genami zgromadzonymi w bazach danych.
SpoÊród oko∏o 30–50 tys. genów ry˝u a˝ 20 tys. jest podobnych do wczeÊniej zbadanych
genów innych organizmów i zak∏ada si´, ˝e pe∏nià one podobne funkcje.
Profilowanie ekspresji genów okreÊla, kiedy i w którym miejscu w roÊlinie gen jest aktywny.
Mikromacierz mieÊci tysiàce fragmentów DNA, tzw. sond, odpowiadajàcych poszczególnym
genom. JeÊli nanieÊç na nià odpowiednio przygotowany ekstrakt komórek, ró˝ne mRNA z
ekstraktu – efekt aktywnoÊci genów – przyczepià si´ do pasujàcych do nich sond, co wi-
doczne jest jako Êwiecenie. AktywnoÊç w okreÊlonym miejscu i czasie – na przyk∏ad wy∏àcz-
nie w fazie rozwoju nasion – oznacza, ˝e gen odgrywa jakàÊ rol´ w tym procesie.
Badany gen mo˝na zniszczyç lub wyciszyç (inaktywowaç) za pomocà odpowiednio skonstru-
owanych fragmentów DNA. Analiza tak zmutowanej roÊliny pod kàtem ró˝nic w budowie lub me-
tabolizmie w stosunku do osobników normalnych mo˝e wskazaç rol´, jakà odgrywa dany gen.
PRZEWIDYWANA KLASYFIKACJA CZYNNOÂCIOWA GENÓW RY˚U
Wykorzystujàc metod´ opisanà powy˝ej, naukowcy ustalili lub przewidzieli funkcje ogrom-
nej cz´Êci genów ry˝u.
Odkryty gen
Baza danych
Podobny gen
Mikromacierz
Próbka
Mutacja
inaktywujàca
gen
RoÊlina
zmutowana
RoÊlina
normalna
AktywnoÊç b∏ony komórkowej
Produkcja energii
Rozwój
Nieznane
Metabolizm
Inne
Transkrypcja
i replikacja DNA
Synteza bia∏ek
Komunikacja komórkowa
Obrona i Êmierç
komórki
Wzrost
i utrzymanie
komórki

pomidory i ry˝, zapoczàtkowali poszu-
kiwania korzystnych alleli genów u
dzikich gatunków, które w przysz∏oÊci
mogà ulepszyç wspó∏czesne roÊliny
uprawne. Ich prace ujawni∏y ogromne
bogactwo genetyczne dzikich krewnych
roÊlin udomowionych i jednoczeÊnie
uÊwiadomi∏y nam, ˝e najcenniejsze za-
soby przyrody sà zazwyczaj na pierw-
szy rzut oka niewidoczne. W jednym z
eksperymentów przeprowadzonych w
po∏owie lat dziewi´çdziesiàtych Tank-
sley i McCouch skrzy˝owali gatunek
drobnego zielonego pomidora z Peru ze
wspó∏czesnà odmianà bladoczerwonà.
Ze zdziwieniem odkryli, ˝e po wprowa-
dzeniu genu z zielonego pomidora bla-
da odmiana sta∏a si´ ciemnoczerwona.
Jak stwierdzono, zielony pomidor nie
mia∏ wprawdzie genów niezb´dnych do
przeprowadzenia syntezy likopenu,
barwnika, który nadaje pomidorom
czerwonà barw´, zawiera∏ jednak „lep-
szy” allel genu warunkujàcego powsta-
nie zwiàzku chemicznego odgrywajàce-
go rol´ na wczeÊniejszym etapie procesu
syntezy likopenu.
Renesans krzy˝owania
DOPIERO ZACZYNAMY
zg∏´biaç genetyczne
zró˝nicowanie dzikich krewnych wspó∏-
czesnych roÊlin uprawnych. Oceniono,
˝e w odniesieniu do ry˝u i pomidora a˝
80% genetycznego zró˝nicowania wy-
st´pujàcego u ich dzikich krewnych a
objawiajàcego si´ ró˝norodnoÊcià alle-
li ka˝dego z genów pozostaje niewyko-
rzystane. Badania Tanksleya, McCouch
i innych wielokrotnie dowiod∏y przydat-
noÊci dzikich alleli do wywo∏ywania zna-
czàcych zmian cech fizycznych u roÊlin
hodowlanych. Zdarza si´, ˝e niektóre z
tych zmian wydajà si´ zachodziç wbrew
oczywistym, widocznym cechom dzi-
kich roÊlin jak w przypadku zielonego
pomidora. Tak wi´c bez technologii wy-
korzystania genów lub loci chromoso-
mowych jako znaczników molekular-
nych naukowcy nigdy nie odkryliby
niektórych po˝àdanych cech ani te˝ nie
wprowadziliby ich do wspó∏czesnych
roÊlin uprawnych.
Skoro zidentyfikowano ju˝ specyficz-
ne grupy korzystnych alleli u ró˝nych
roÊlin dzikich lub uprawnych, nale˝a-
∏oby przenieÊç je (i tylko je) do szcze-
pów szlachetnych odmian uprawnych.
Do tego celu mo˝na wykorzystaç bak-
teryjny DNA albo jakikolwiek inny Êro-
dek transferu wybranych genów, sto-
sujàc proces zwany transformacjà, cha-
rakterystyczny dla metod tworzenia
˝ywnoÊci modyfikowanej genetycznie.
Jednak aby uniknàç d∏ugotrwa∏ej i kosz-
townej procedury dopuszczenia trans-
genicznych roÊlin do sprzeda˝y, naukow-
cy próbujà tak˝e techniki tradycyjnego
krzy˝owania ukierunkowanego przez
zastosowanie znaczników genetycznych.
Znajàc dok∏adnie allele niosàce po˝à-
dane cechy, albo przynajmniej ich loka-
lizacje chromosomowe, hodowca po-
trafi „zaprojektowaç” roÊlin´ ∏àczàcà te
nowe cechy z najkorzystniejszymi w∏a-
ÊciwoÊciami szlachetnych odmian upraw-
nych. RoÊlin´ takà mo˝na nast´pnie
wyhodowaç drogà zaplanowanych krzy-
˝ówek wspomaganych technikà znacz-
ników DNA (pomocnà np. przy ustalaniu
ojcostwa i w medycynie sàdowej), dzi´ki
której selekcjonerzy mogà sprawdziç
obecnoÊç po˝àdanych genów w nowych
krzy˝ówkach [ramka na nast´pnej stro-
nie]. Jak we wszystkich wielkoskalowych
hodowlach roÊlin ka˝da taka krzy˝ówka
daje dziesiàtki tysi´cy roÊlin potomnych.
Ale zamiast hodowania ka˝dej z nich i
d∏ugiego oczekiwania na pe∏ny rozwój,
by si´ przekonaç, którym osobnikom zo-
sta∏a przekazana korzystna cecha, ho-
dowca móg∏by po prostu pobraç ma∏à
próbk´ DNA z ka˝dej siewki i sprawdziç
jej geny na obecnoÊç alleli – znaczników
po˝àdanych cech.
Siewki majàce po˝àdane cechy ros∏y-
by nast´pnie do momentu osiàgni´cia
gotowoÊci do skrzy˝owania z wyjÊciowà
odmianà uprawnà. Ich potomstwo by-
∏oby dalej testowane pod kàtem obecno-
Êci tego specyficznego allelu a˝ do czasu,
gdy hodowca otrzyma∏by populacj´ ro-
Êlin przypominajàcych wyjÊciowà od-
mian´ uprawnà, ale zawierajàcà nowo
wprowadzony allel warunkujàcy korzyst-
nà cech´. Dzi´ki wykorzystaniu znacz-
ników genetycznych do potwierdzania
obecnoÊci po˝àdanych cech na ka˝dym
etapie tego procesu mo˝na by o wiele lat
skróciç okres niezb´dny do wyhodowa-
nia nowych odmian uprawnych. To po-
zwoli∏oby selekcjonerom szybciej reago-
waç na zmiany warunków upraw, na
przyk∏ad pojawienie si´ nowych szkod-
ników lub ich uodpornienie na dotych-
czasowe metody zwalczania. ¸atwiejsze
by∏oby tak˝e dopasowanie nowych od-
WRZESIE¡ 2004 ÂWIAT NAUKI
37
KA
Y CHERNUSH
STEPHEN A. GOFF i JOHN M. SALMERON sà genetykami roÊlin w Syngenta Biotechnology,
Inc. w Research Triangle Park w Karolinie Pó∏nocnej. Goff kierowa∏ amerykaƒskim zespo∏em,
który w 2002 roku zsekwencjonowa∏ genom ry˝u. Obecnie w ramach akcji humanitarnej pracu-
je na rzecz wykorzystania tej wiedzy do poprawy plonowania zbó˝ w krajach rozwijajàcych si´.
Salmeron, dyrektor dzia∏u genetyki stosowanej w SBI, wprowadza∏ genetyk´ do procesu udosko-
nalania zbó˝ ju˝ od 1989 roku, gdy po doktoracie jako stypendysta University of California w
Berkeley wyizolowa∏ z pomidora jeden z pierwszych roÊlinnych genów odpornoÊci na choroby.
O
AUTORACH
AUTORZY, JOHN SALMERON (z lewej) I STEPHEN GOFF, z doÊwiadczalnymi roÊlinami kuku-
rydzy. W szklarni kwiatostany ˝eƒskie dojrza∏ych do rozmna˝ania si´ roÊlin kukurydzy nakry-
wane sà ma∏ymi torebkami z pergaminu, co zapobiega ich zapyleniu przez zlokalizowane na
szczycie roÊlin kwiatostany m´skie, przypominajàce fr´dzle.
38
ÂWIAT NAUKI WRZESIE¡ 2004
SLIM FILMS (
ilustracje
); RICE PHOTOGRAPHS PPRZEDRUK ZA ZGODÑ T
ANKSLEY
A I MCCOUCH Z
SCIENCE
, TOM 277; 1997. ©
1997
AAAS; C. MAR
T
INEZ
International Center for T
ropical Agricultur
e
(z lewej
); CORNELL UNIVERSITY (
z prawej
)
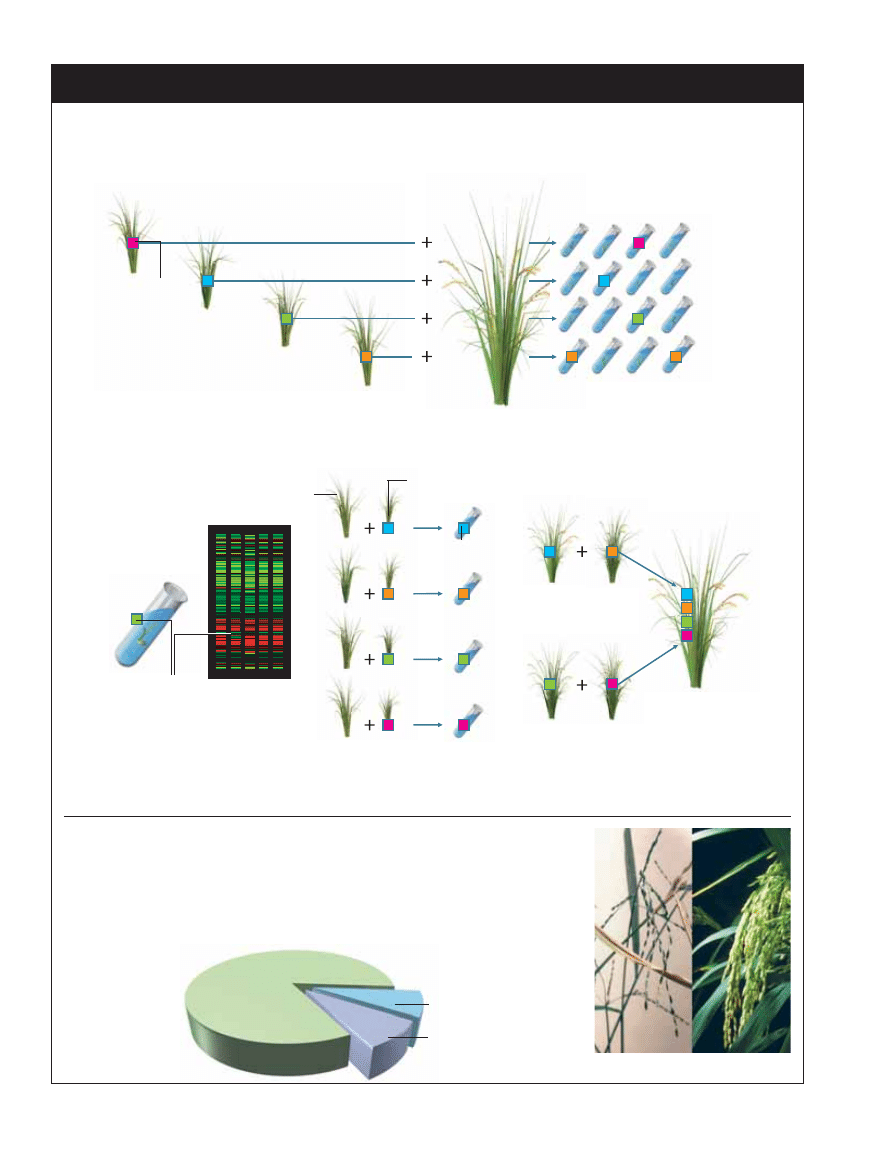
TWORZENIE NOWYCH ODMIAN ROÂLIN
GDY JU˚ WIADOMO, który gen kontroluje specyficznà cech´ roÊliny (np. wielkoÊç nasion), mo˝na badaç ró˝ne odmiany roÊlin uprawnych
i ich dzikich krewnych w poszukiwaniu wariantu genu (allelu) korzystnie kszta∏tujàcego t´ cech´. Znaleziony allel przenosi si´ z jednej
roÊliny do drugiej metodà konwencjonalnego krzy˝owania. Tradycyjna selekcja wymaga nast´pnie hodowania uzyskanych roÊlin do
czasu, gdy korzystna cecha si´ ujawni. ZnajomoÊç allelu pozwala na zastosowanie go jako genetycznego znacznika identyfikujàcego skutecznie
skrzy˝owane roÊliny zaraz po ich wykie∏kowaniu, co skraca czas wytwarzania nowych odmian.
1
Ka˝da z czterech ró˝nych odmian ry˝u z po˝àdanà cechà (reprezentowanà
przez barwne kwadraty) mo˝e byç krzy˝owana ze szlachetnà odmianà
uprawnà, dajàc dziesiàtki tysi´cy siewek.
2
Tylko niektóre siewki b´dà zawie-
raç rodzicielski allel genu, warun-
kujàcy po˝àdanà cech´.
3
Zamiast hodowaç tysiàce roÊlin a˝ do cza-
su pojawienia si´ po˝àdanej cechy, ju˝ w
kilka dni po wykie∏kowaniu mo˝na technikà
genetycznego „odcisku palca” testowaç obec-
noÊç korzystnego allelu w ka˝dej siewce.
4
Tylko roÊliny potomne z korzystny-
mi allelami sà uprawiane dalej. Gdy
dojrzejà, krzy˝uje si´ je z wyjÊciowà od-
mianà uprawnà (krzy˝owanie wsteczne).
5
Krzy˝owanie, krzy˝owanie wsteczne i te-
stowanie DNA jest powtarzane, dopóki
wszystkie korzystne allele nie zostanà prze-
niesione do szlachetnej odmiany uprawnej.
RÓ˚NORODNOÂå GENETYCZNA RY˚U
Po tysiàcach lat krzy˝owania wsobnego (w grupie blisko spokrewnionych osobników) ró˝norodnoÊç
genetyczna roÊlin uprawnych jest znacznie mniejsza ni˝ ich dzikich krewnych, które stanowià bogaty
„magazyn” nowych, cennych alleli. Zalety si´gania do tych zasobów nie zawsze sà oczywiste. W jednym
z doÊwiadczeƒ allele dzikiego przodka ry˝u Oryza rufipogon (z lewej) przeniesiono do wspó∏czesnej
wysokoplennej odmiany chiƒskiego ry˝u (z prawej) za pomocà krzy˝owania wspomaganego znacznikami
DNA. Geny s∏abo plonujàcej dzikiej roÊliny podnios∏y plonowanie nowoczesnej odmiany o 17–18%.
Odmiany dzikie
lub egzotyczne
O. sativa indica
O. sativa japonica
Po˝àdana
cecha
Ró˝ne odmiany ry˝u
Szlachetna
odmiana uprawna
Siewki
Odmiana
szlachetna
z po˝àdanymi
allelami
Ulepszona
odmiana
szlachetna
Siewki
z krzy˝owania
wstecznego
Szlachetna
odmiana uprawna
Potomstwo z po˝àdanymi allelami
Po˝àdany
allel
Genetyczny
„odcisk palca”
O. rufipogon
Odmiana dajàca
wysokie plony
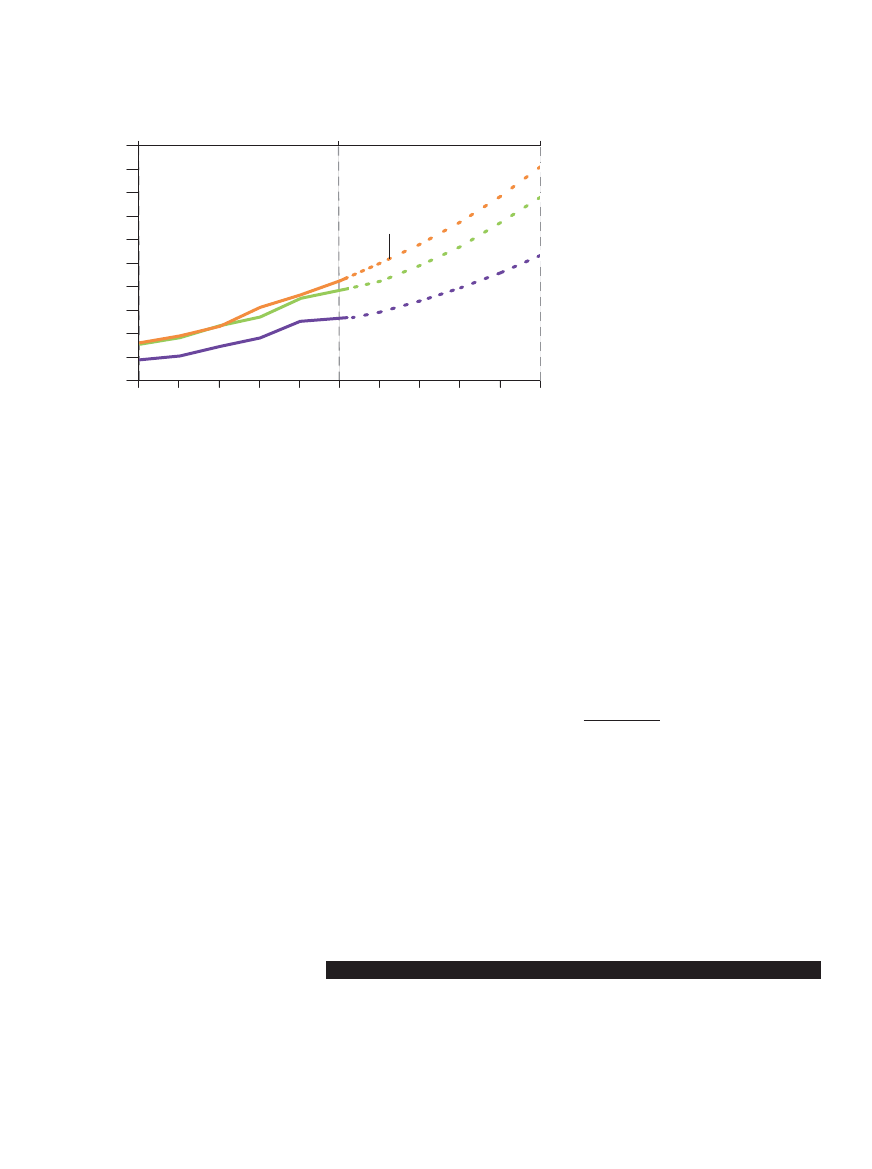
mian uprawnych do najró˝niejszych wa-
runków Êrodowiskowych, potrzeb rolni-
ków i preferencji konsumentów.
Prawdziwa rewolucyjnoÊç tej meto-
dy polega jednak na mo˝liwoÊci posze-
rzenia genetycznego „wàskiego gard∏a”
powsta∏ego tysiàce lat temu, gdy udomo-
wiono najwa˝niejsze roÊliny. Zgroma-
dzenie danych na temat funkcji genów
traw pozwoli efektywniej przeszukiwaç
ten ogromny magazyn ró˝norodnoÊci
genetycznej. Przyk∏adem takich mo˝li-
woÊci jest jedno z doÊwiadczeƒ zespo-
∏u McCouch, w którym wykorzystano
znaczniki molekularne do zidentyfiko-
wania loci genów w dzikim przodku ry-
˝u Oryza rufipogon, rosnàcym do dzi-
siaj w Malezji, a nast´pnie zastosowano
wspomaganà znacznikami hodowl´ w
celu przeniesienia 2 tys. genów – oko∏o
5% genomu ry˝u – z dzikiego gatunku
do mieszaƒca wspó∏czesnych odmian
ry˝u chiƒskiego.
Celem eksperymentu by∏o odnalezie-
nie alleli umo˝liwiajàcych dalsze zwi´k-
szenie ju˝ i tak wysokiej produktywnoÊci
tego mieszaƒca. Nowo powsta∏e roÊliny
testowane by∏y na obecnoÊç kilku cech
zwi´kszajàcych plonowanie, takich jak
wysokoÊç roÊliny, d∏ugoÊç k∏osa, masa
ziaren. Oko∏o po∏owa loci pochodzàcych
od dzikiego krewnego zawiera∏a allele
poprawiajàce plonowanie, jednak nie-
które z nich mia∏y negatywny wp∏yw na
wzrost roÊlin, na przyk∏ad opóênia∏y doj-
rzewanie. Jednak˝e dwa z alleli wyst´-
pujàcych w O. rufipogon wydawa∏y si´
nie mieç ˝adnego negatywnego wp∏ywu
i spowodowa∏y wzrost plonów o 17–18%
w porównaniu z nowoczesnà odmianà.
Tak jak w eksperymencie McCouch z zie-
lonym pomidorem nic w wyglàdzie dzi-
kiej roÊliny [ramka na poprzedniej stronie]
nie zapowiada∏o, ˝e mo˝e si´ ona przy-
czyniç do wzrostu plonowania nowocze-
snej odmiany. Tym bardziej wi´c rezulta-
ty by∏y zach´cajàce.
OczywiÊcie nie wszystkie korzystne ge-
ny mogà byç przeniesione do wspó∏cze-
snych odmian drogà tradycyjnej hodow-
li. Na przyk∏ad genów warunkujàcych
tolerancj´ na herbicydy lub niektóre typy
odpornoÊci na owady nie ma w roÊlinach,
które mogà krzy˝owaç si´ z kukurydzà.
W takich przypadkach po˝àdany gen mo-
˝e zostaç przeniesiony do docelowej ro-
Êliny technikà transformacji za pomocà
DNA bakterii lub innego noÊnika, nie jest
jednak wtedy mo˝liwe przewidywanie i
kontrolowanie miejsca, w którym nowy
gen lokuje si´ w genomie roÊliny biorcy.
Mo˝na wi´c dodaç nowy allel, ale nieko-
nieczne zastàpiç nim stary, mniej po˝àda-
ny. Natomiast podczas modyfikacji ssa-
ków lub niektórych drobnoustrojów, dzi´-
ki zjawisku nazywanemu rekombinacjà
homologicznà, wprowadzany gen jest
kierowany do miejsca w chromosomie,
którego sekwencja DNA jest najbardziej
do niego zbli˝ona, pozwalajàc na precy-
zyjne zastàpienie genu oryginalnego
przez po˝àdany allel. Mo˝e uda si´ nam
w przysz∏oÊci osiàgnàç takie samo jed-
noetapowe zast´powanie alleli genów w
roÊlinach uprawnych. Ostatnio zastoso-
wano rekombinacj´ homologicznà do
modyfikowania ry˝u, a zbli˝ony proces
wykorzystano do wymiany alleli w ku-
kurydzy. Gdy stanie si´ to rutynà, umie-
j´tnoÊç zast´powania w ten sposób od-
cinków chromosomów w warunkach
laboratoryjnych umo˝liwi naukowcom
przenoszenie genów mi´dzy roÊlinami,
które nie dajà si´ krzy˝owaç w sposób
naturalny
2
.
Genetyczne wspomaganie krzy˝owa-
nia skraca procedur´ wyprowadzania
nowych odmian u roÊlin tego samego
gatunku lub blisko spokrewnionych. W
ciàgu ostatnich 3 tys. lat nie udomowio-
no ˝adnych nowych gatunków zbó˝. Tak
wi´c aby zaspokoiç stale rosnàce zapo-
trzebowanie na ˝ywnoÊç, b´dziemy mu-
sieli skoncentrowaç si´ na udoskona-
laniu istniejàcych najwa˝niejszych roÊlin
uprawnych. Nauki genetyczne, dostar-
czajàc narz´dzi do przeszukiwania ge-
nomów i wykorzystywania ukrytych
w nich skarbów, rozpoczynajà nowà
zielonà rewolucj´.
n
1
Ziarna zbó˝ sà w rzeczywistoÊci owocami nale-
˝àcymi do grupy ziarniaków, a nie – jak si´ mo˝e
wydawaç – nasionami. Poniewa˝ tematem artyku-
∏u sà zbo˝a, utrzymano termin „ziarna”, choç opi-
sane mechanizmy równie dobrze odnoszà si´ do
hodowli i selekcji innych roÊlin uprawnych.
2
Warto przy okazji zauwa˝yç, jak z∏udne sà proste
podzia∏y roÊlin uprawnych na otrzymane „natural-
nie” oraz transgeniczne (czyli genetycznie zmody-
fikowane – GMO). Przeniesienie cennego allelu dro-
gà tradycyjnego krzy˝owania uprawnia roÊlin´ do
uzyskania statusu „naturalnego mieszaƒca”. Na-
tomiast przeniesienie tego samego allelu drogà ho-
mologicznej rekombinacji kwalifikuje ju˝ roÊlin´
do kategorii GMO! Z drugiej strony z kategorii GMO
wy∏àczono organizmy otrzymane drogà losowej
mutagenezy wywo∏anej dzia∏aniem wysokich da-
wek promieniowania jonizujàcego lub substancji
mutagennych.
WRZESIE¡ 2004 ÂWIAT NAUKI
39
NINA FINKEL: èRÓD¸
O: FOOD AND AGRICUL
TURE ASSOCIA
T
ION
Early Allelic Selection in Maize as Revealed by Ancient DNA. Viviane Jaenicke-Després i in.;
Science, tom 302, s. 1206-1208; 14 XI 2003.
Seed Banks and Molecular Maps: Unlocking Genetic Potential from the Wild. Steven D. Tanksley
i Susan R. McCouch; Science, tom 277, s. 1063-1066; 22 VIII 1997.
A Draft Sequence of the Rice Genome (Oryza sativa L. ssp. japonica). Stephen A. Goff i in.;
Science, tom 296, s. 92-100; 5 IV 2002.
Gramene, a Tool for Grass Genomics. D. H. Ware i in.; Plant Physiology, tom 130, nr 4, s. 1606-
-1613; XII/2002.
JEÂLI CHCESZ WIEDZIEå WI¢CEJ
0
2000
4000
6000
8000
10 000
1950
1960
1970
1980
1990
2000
2010
2020
2030
2040
2050
Lata
Kilogramy na hektar
WZROST PLONOWANIA ZBÓ˚
Pszenica
Kukurydza
Ry˝
2.5 mld
6 mld
9 mld
LudnoÊç Êwiata
Prognozowane potrzeby
ÂREDNIE PLONY z pszenicy, kukurydzy i ry˝u na Êwiecie zwi´kszy∏y si´ niemal trzykrotnie w la-
tach 1950–2000, w tym samym czasie podobnie wzros∏a ludnoÊç Êwiata. Aby w 2050 roku wy-
karmiç przewidywane 9 mld ludzi przy zachowaniu obecnego dziennego spo˝ycia zbó˝ na po-
ziomie 0.4–1.5 kg na osob´, plony powinny wzrastaç co roku o 1.5%.
Wyszukiwarka
Podobne podstrony:
3786
200409 3784
200409 3777
pd5 e lubiszewska 20040903
200409 3795
3786
200409 3776
200409 3782
200409 3785
200409 3767
200409 3796
200409 3790
200409 3788
200409 3798
200409 3791
200409 3794
200409 3768
więcej podobnych podstron