DSCN6176 (Kopiowanie)

159
4 Struktura mikroskopowa organizmów żywych; komórki I tkanki bezpośrednio z jednej komórki na drugą. Synapsy elektryczne są rzadkie, występująjcdynie w ośrodkowym układzie nerwowym; mogą one przewodzić impulsy dwukierunkowo i nic podlegają zmęczeniu.
Synapsy chemiczne znacznie bardziej rozpowszechnione - są większe i mają szerszą szczelinę (ok. 20 nm). Impuls przez synapsę przechodzi wyłącznic od ncurytu jednej komórki do dendrytu drugiej - nigdy odwrotnie. Dzieje się tak dzięki sckrecyjncj czyli wydziclniczcj zdolności komórki nerwowej. Zakończenie aksonu stanowi kolbka synaptyczna (ryc. 4-31), w której obecne są mitochondria, a przede wszystkim pęcherzyki synaptyczne zawierające mediator.
Większość mediatorów stanowią substancje chemiczne znane już wcześniej jako hormony. Impuls docierający do kolbki synaptycznej sprawia, że pęcherzyki łączą się z błona prcsvnanlvczna i uwalniają swą zawartość do szpary synaptycznej. Mediator łączy się wówczas z błona postsvnaptvczna będącą fragmentem dendrytu drugiej komórki i powoduje powstanie w niej impulsu. Po czym cząsteczka mediatora zostaje natychmiast rozłożona przez odpowiednie enzymy obecne w szczelinie synaptycznej.
Dziś wiemy, że neurony mogą uwalniać wytworzone przez siebie mediatory również do krwi, gdzie działają one jako hormony. Zjawisko takie nosi nazwę ncurosekrecii. Każda ze struktur narysowanych na ryc. 4-32 może być zakończeniem neuronu ncurosekrecyjnego albo kolbką synaptyczną łączącą dwie komórki nerwowe. Neuron cholincmczny produkuje mediatoracetylocholine, neuron adrcncrgiczny -adrenalinę, noradrenalinę bądź dopamine (omówione zostaną w rozdziale o hormonach), zaś neuron peptvdrcgicznv mediatory o budowie białkowej (w ich transporcie ważną rolę odgrywają wlókicnka zwane ncurofibrylami). Oprócz opisanych powyżej synaps chemicznych istnieją także połączenia, w których z jednej komórki do drugiej przepływająjony (np. wapniowe, sodowe, potasowe). Połączenia te nazywamy synapsami jonowymi. Występują one w tkance nerwowej, ale też i w innych tkankach (np. połączenia szczelinowe, takie jak omówione już wstawki w mięśniu sercowym).
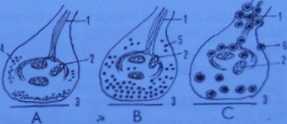
Ryc. 4 - 32. Ultrastrukturalne modele zakończeń neuronów wydzielniczych: A - cholincrgiczncgo; B - adrcncrgiczncgo;
C - pcptydergiczncgo (JD).
I - ncurotubulc; 2 - mitochondria;
3 - szpara synaptyczna;
4 - pęcherzyki synaptyczne zawierające acetylocholinę:
5 - ziarna wydziclniczc typu B o średnicy do 100 nm. zawierające
aminy biogenne:
6- ziarna wydziełnicze typu A o średnicy 200nm. zawierające peptydy.
4.5. Tkanki roślinne
Wprawdzie prototypy tkanek występują w rozwoju ewolucyjnym już u mszaków (skórka, tkanka miękiszowa i prymitywne elementy przewodzące w łodydze) i u paprotników (tkanka naczyniowa), jednak wszystkie opisane poniżej tkanki roślinne obserwować możemy dopiero u roślin naczyniowych. Dzielimy je na dwie duże grupy: na tkanki twórcze i tkanki stałe.
4.5.1. Tkanki twórcze
Tkanki twórcze czyli merystemy składają się z małych komórek o dużych jądrach i cienkich ścianach. Zawierają dużo cytoplazmy i nie posiadają wodniczek (bądź też mająje nieduże i nieliczne). Funkcją tych komórek jest dzielenie się (ryc. 4-20) i różnicowanie, w następstwie którego powstają tkanki wszystkich innych typów.
Merystemy z punktu widzenia ich powstania możemy podzielić na pierwotne i wtórne. Pierw otne powstają przez podziały zygoty i tworzą początkowo rozwijający się wielokomórkowy zarodek. W miarę rozwoju większość tych komórek ulega zróżnicowaniu na tkanki stale, lecz nawet w starych roślinach w pewnych miejscach pozostaje tkanka twórcza zapewniająca nieprzerwany wzrost (wierzchołki korzeni ■pędów, miazga wiązek naczyniowo-sitowych).
Wyszukiwarka
Podobne podstrony:
DSCN6162 (Kopiowanie) 4 Strukturo mikroskopowa organizmów żywych: komórki I tkanki Tabela 4-IV. Poró
DSCN6164 (Kopiowanie) V 4 Struktura mikroskopowa organizmów żywych: komórki i tkanki 147 (śródblonc
DSCN6164 (Kopiowanie) 4 ^Struktura mikroskopowa organizmów żywych: komórki i tkanki 147 4 ^Struktura
DSCN6152 (Kopiowanie) I3S 4. Stmknm mikrmkopona organizmów żywych: komórki i tkanki U pierwotniaków
DSCN6166 (Kopiowanie) ’f, Snukmm mikroskopowa organizmów iywych: komórki i tkanki 149 ’f, Snukmm mik
DSCN6176 (Kopiowanie) 159 i mikroskopowa organizmów żywych: komórki i tkanki bezpośrednio z jednej k
DSCN6150 (Kopiowanie) 4 Stmkrura mikroskopowa organizmów żywych: komórki i tkanki 33 T faraoniv opis
DSCN6166 (Kopiowanie) I 4, Stmktura mikroskopowa organizmów żywych: komórki i tkanki 4.4.1.3. Tkanka
DSCN6174 (Kopiowanie) mikroskopowa organizmów żywych: komórki i tkanki 157 mikroskopowa organizmów ż
DSCN6142 (Kopiowanie) 125 4. Srnhm mikroskopowa aryanUmmr tywyrk. komórki I tkanki Zawiesinę możemy
DSCN6140 (Kopiowanie) mikn»kopow<i organizmów żywych: komórki i tkanki Isl 123 r^bek szczote
DSCN6160 (Kopiowanie) I 4 StnĄtum mikroskopowi/ organizmów śywyćfi: komórki i Ikanki ł 143 I 4
DSCN6162 (Kopiowanie) ^ £‘truktura mikroskopowi organizmów ^yiyych: komórki i tkanki 145 ^ £‘truktur
DSCN6170 (Kopiowanie) tofryktum miKrosmtptmu urgunJzftiow żywych: komórki i
DSCN6112 (Kopiowanie) 97 ł Sirfkiuni molekularni! istot żywych3. STRUKTURA MOLEKULARNA ISTOT ŻYWYCH
DSCN6115 (Kopiowanie) 99 V I jyebwiw molekularna istot żywych ■fi8ft!,adu ma^Uniąją j Sązaien k
DSCN6118 (Kopiowanie) 101 ł StniMura molekularna ittot żywych -$5 lnej
DSCN6135 (Kopiowanie) X d = 0,61 -A (2) obiektywu mikroskopu. mikroskop do obserwacji i badań naukow
DSCN6157 (Kopiowanie) W jąderku obserwowanym w mikroskopie elektronowym wyróżnić możemy dwie części:
więcej podobnych podstron